|
||
|
|
||
| Главная ≫ Инфотека ≫ Разное ≫ Книги ≫ Глава первая. Эволюция. Возникновение сложности / Десять великих идей науки. Как устроен наш мир // Питер Эткинз |
Глава первая. Эволюция. Возникновение сложности / Десять великих идей науки. Как устроен наш мирПитер Эткинз
Глава первая. Эволюция. Возникновение сложностиБез света эволюции ничто в биологии не имеет смысла. Великая идея: эволюция идет путем естественного отбораЖизнь столь совершенна, что, как долгое время считали, ее было необходимо сотворить особо. Ибо как может нечто столь удивительное и столь уникальное самопроизвольно возникнуть из безжизненной слизи? В самом деле, что является той главной частью вещей, которая наделяет их жизнью? Ответы на эти и другие вопросы первостепенной важности появились двумя волнами. Первой была волна эмпирических объяснений, когда наблюдатели, по большей части натуралисты и геологи девятнадцатого века, пристально изучали внешние формы природы и получали далеко идущие выводы. Затем пришла вторая волна, в двадцатом веке, когда кроты с глазами ученых рыли ходы под поверхностью явлений и открывали молекулярную основу паутины жизни. Первый из этих подходов является предметом настоящей главы; второй, чрезвычайно обогативший наше понимание того, что значит быть живым, есть предмет следующей.Древнегреческие философы, как обычно, имели свои собственные взгляды на природу живых существ. Как и большинство их мнений, высказанных из самых лучших побуждений, эти взгляды были нелепо, но обаятельно превратны. Например, самопровозглашенный бог Эмпедокл (490-430 до н.э.), незадолго до принятия им не слишком мудрого решения продемонстрировать свою божественность путем низвержения себя в кратер вулкана Этна, предположил, что животные построены из универсального набора частей, которые, будучи собраны в различные комбинации, дают слона, комара, рогатую жабу и человека. Мир заселяется этими знакомыми комбинациями охотнее, чем летающими свиньями и ослами с рыбьим хвостом потому, что лишь некоторые комбинации жизнеспособны. Природа, вероятно, экспериментировала с другими комбинациями, в ожидании Острова доктора Моро, но после недолгого хромания, трепыхания и переваливания с боку на бок эти экспериментальные творения умирали. Почти на два тысячелетия позже эхом откликнулся похожий взгляд, но уже на молекулярном уровне. Его высказал граф Жорж-Луи Леклерк де Бюффон (1707-88), считавший, что организмы самопроизвольно возникли из агрегаций, которые мы сегодня назвали бы органическими молекулами, и что число возможных видов есть число жизнеспособных комбинаций этих молекул. Бюффон полагал, что уж он-то знает: свой великий труд Всеобщая и частная естественная история (Histoire naturelle, générale, et particulière), начатый в 1749 г., он планировал довести до пятидесяти томов, но успел подготовить тридцать шесть. Девять были посвящены птицам, пять минералам и восемь (опубликованных посмертно) китообразным, рептилиям и рыбам. Но откуда все эти творения, все живые существа действительно появились, их внушающее благоговение количество, два миллиона зарегистрированных видов и, возможно, десять и более миллионов, которые еще предстоит обнаружить? Аристотель, всегда блистательно обильный интеллектуально и, как всегда, блистательно ошибавшийся, предположил, что животные упали со звезд или произошли самопроизвольно уже в завершенном виде. Индейцы яхуна из бассейна Амазонки приняли неоаристотелевский взгляд и считали, что маниока выросла из пепла убитого и кремированного Миломаки. Индейцы кауилла из Калифорнии также верили, что виноградная лоза выросла из его кремированного живота, арбузы из зрачков его глаз, а злаки из его зубов. Менее общепринято мнение, что пшеница произошла из яиц его вшей, а бобы из его спермы. Другие религии предложили с виду простые мнения, согласно которым все существа, большие и малые, были сотворены Богом, и это все. Однако даже некоторые отцы церкви находили затруднительным примириться со всеми утверждениями Библии. Например, эрудит Григорий Назианин (330-89, Назиан был где-то в Каппадокии, в Малой Азии), считал, что Бог должен был создать некоторые из своих творений после потопа, поскольку небольшой ковчег Ноя был слишком мал для того, чтобы приютить представителей всех видов.[1] Архидиакон Карлайла Уильям Пэйли считал бесспорным, что он установил происхождение творений в книге с игривым названием Естественная теология или свидетельства существования и атрибуты божества, собранные из природных явлений, опубликованной в 1802 г., где в качестве превосходного аргумента он использует аналогию с путешественником, который столкнулся с ручными часами, созерцает их сложный замысел и не сомневается в том, что за этим замыслом стоит часовщик. Таким образом, каждый, кто сталкивается со сложностью природы должен неизбежно заключить, что к ее замыслу и конструкции приложил руку Бог. Однако Анаксимандр из Милета (610-545 до н.э.), делая вклад в западную философию, когда она была еще едва пробившимся зеленым побегом, действительно имел проблеск чего-то, похожего на истину. В совместной с Фалесом и Анаксименом философской программе он сделал чисто умозрительное заключение относительно всех живых существ и жизни в целом, гласящее, что виды животных могут превращаться один в другой. В науке часто бывает, что первым шагом к действительному пониманию, приходящему на смену фантастическим спекуляциям, оказывается собирание данных. В данном случае для этого надо идентифицировать и классифицировать все типы организмов, составляющих биосферу, или, по крайней мере, столько, сколько позволят терпение, настойчивость и провидение. Наиболее полезными бывают названия, выражающие родство, похожие на обычай давать членам одной семьи одну фамилию. К середине восемнадцатого века, когда установилась международная морская торговля, даже домоседы стали осведомленными об изобилии организмов и странных существ, населяющих мир, и поняли, что простых названий, таких как корова и собака, уже недостаточно. Это было подобно тому, как обитатели Лапландии вдруг обнаружили бы, что в Уганде недостаточно знания их родного языка. Первую общепризнанную систему наименований разработал шведский ботаник Карл фон Линней (Carl von Linne, 1707-78), в латинском написании Линнеус (Linnaeus). Линней изложил свою систему наименований в труде Systema naturae, опубликованном в 1735 г., а для извлечения на свет систематической классификации растений обычно обращаются к его книге Spesies plantarum, 1753. В этих работах Линней ввел иерархию групп (рис. 1.1) с царствами около вершины и пирамидой, расширяющейся ко все более частным разделам по мере нисхождения через типы, классы, отряды, семейства, роды и виды. Эта схема с тех пор пополнялась посредством включения различных промежуточных слоев, таких как подсемейство и надсемейство. Так, нас, людей, следует классифицировать (иронически — возразит кто-то) как вид Homo sapiens, рода Homo, в семействе Гоминидов, в надсемействе Гоминоидов инфраотряда Узконосых подотряда Антропоидов отряда Приматов подкласса Плацентарных в классе Млекопитающих, в надклассе Четвероногих из типа Хордовых в царстве Животных домена Ядерных в империи организмов.
Недостатком системы Линнея является то, что она основана на внешне узнаваемых подобиях скорее, чем на более приемлемом в научном отношении установлении лежащего в их основе родства. Более того, точному определению классов, типов и т. д. трудно приписать, а в действительности и вовсе не удается какой-либо особенно глубокий фундаментальный смысл. Современным методом таксономии является кладистика (кладос, по-гречески молодой побег), тщательно исследующая происхождение организмов от общего предка и идентифицирующая различные ветви, или клады, дерева жизни (рис. 1.2). Кладйстика была введена немецким таксономистом Вилли Хеннигом (1913-1976) и доработана в его Филогенетической систематике (1966). Согласно Хеннигу, классификация должна отражать генеалогическое родство, и организмы должны группироваться строго на основе их происхождения от общего предка. Не в пример легкомысленным теоретическим физикам, приспособившим к своим схемам повседневные слова, такие как «спин» (в переводе с английского «вращение») и «аромат», Хенниг перегрузил таксономию греческими терминами, и кладисты имеют дело с симплециоморфами (характеристиками, разделяемыми более чем одной тварью), синапоморфами (разделяемыми производными характеристиками) и т.д. К счастью, у нас нет необходимости использовать этот перегруженный язык, поскольку мы главным образом будем пользоваться системой Линнея. Однако кладистика весьма влиятельна, логична и полезна, так как она основана на генеалогии организмов, которая, как можно доказать, есть единственный рациональный фундамент классификации.
Немедленно, однако, мы натыкаемся на сложную проблему, которая пронизывает все дальнейшее обсуждение и служит предметом беспокойства даже для новейших систем классификации: какое содержание мы вкладываем в термин «виды». Даже сегодня нередко ведутся споры о его точном определении. Такие споры не имеют большого практического значения, однако, поскольку эта концепция является центральной в исторической дискуссии о происхождении видов, необходимо, по крайней мере вкратце, ее коснуться. Может быть, в действительности было бы лучше признать невозможность изобретения универсально пригодного определения, считая термин «виды» внутренне неопределенным и не возводя излишне жестких стен ради его фиксации. У тех, кого иногда называют типологическими таксономистами, обычно принято определять вид как группу организмов, которые выглядят — по опознаваемым морфологическим признакам — отличными от организмов из других групп. Платон воспользовался весьма похожей идеей в своей концепции эйдоса, или «совершенной формы», идеала, истинной сущности, лишь несовершенно представляемой реальными существами. Мы без труда отличаем воробья от черного дрозда по их «идентифицируемым морфологическим признакам» и считаем их разными видами птиц. Для нас, как мы полагаем, не составляет труда опознать сущностную «птичность» этих двух творений и усмотреть ее отличие от «растениевости» репы, а также отличить «воробьевость» одного от «чернодроздовости» другого. Несколько более изощренное определение имеет концепция биологического вида, определяющая вид как группу организмов, которые спариваются между собой, но репродуктивно изолированы от других подобных групп. Согласно этому взгляду, вид является изолированным островом с повышенной репродуктивной активностью. Такое определение помещает воробья и черного дрозда в разные виды, поскольку они размножаются каждый внутри своей группы и не спариваются между собой. Репродуктивная изоляция может возникать различными путями. Например, группы организмов могут быть географически изолированы — это одна из причин, делающих острова столь важными в истории эволюционных идей — или размножаться в разные времена года. Группы могут находить друг друга отталкивающими (или, по крайней мере, лишенными привлекательности) или обнаружить травмирующую физическую невозможность спаривания, как бы их ни влекло друг к другу. Предвкушая механизм наследственности, речь о котором пойдет в следующей главе, мы могли бы сказать, что каждый вид представляет собой отдельный генный бассейн, с генами, циркулирующими внутри бассейна при спаривании представителей вида — процесс, называемый генным потоком, — но не мигрирующими в генные бассейны, представляющие другие виды. Генный поток внутри вида гарантирует, что все его представители выглядят более или менее похожими, так что концепция биологических видов согласуется с критериями, принятыми у типологических таксономистов. Тогда почему же определение видов столь противоречиво? Одна из проблем в определении, основанном на понятии спаривания, состоит в том, что некоторые организмы вообще не спариваются. Например, далеко не все спаривающиеся бактерии классифицированы как виды, и существует множество примеров многоклеточных организмов, которые размножаются неполовым путем (таких, как обычные одуванчики, Taraxacum officinale), однако считаются истинными видами. Эта проблема обнаруживает, что слово «вид» имеет два, иногда несхожих, значения. Одно значение, отсылающее к сказанному выше, связано с репродуктивной изоляцией организмов. Второе значение состоит в том, что термин «вид» является просто одной из конечных точек в основании таксономической пирамиды, предельной единицей классификации групп организмов, безотносительно к их способности или неспособности спариваться с другими организмами. То есть вид это в точности таксон, единица классификации. Использование термина «вид» просто для обозначения таксона является обычным в палеонтологии, где одной линии наследования могут приписываться различные имена на разных стадиях ее развития, хотя ее последовательные члены никогда не могли бы даже рассмотреть возможность спаривания. Так, Homo erectus превратился в H. sapiens, но они никогда не прогуливались вместе: оба являются примерами того, что иногда называют хронотипами. Осознание этих трудностей создает мотив для поиска альтернативных определений вида, которые то перекрываются, то вступают в конфликт с биологической концепцией видов. Например, один из способов классификации организмов является фенетическим, в нем организмы зачисляются в одну группу на основании чисто объективных измерений, включая дискретные измерения, такие как использование числа 1 для факта «имеет крылья» и 0 для «не имеет крыльев». Игры «узнай своего партнера» в газетах, журналах и агентствах по сбору данных являются по сути фенетическими. Преимущество фенетического подхода в том, что он строго объективен и не полагается на субъективные суждения о происхождении организмов и на догадки о том, могли, при случае, один организм — возможно уже исчезнувший — спариться с другим. Одна из проблем этой схемы состоит в том, что хотя фенетически отождествляемые группы организмов выглядят почти идентичными, они тем не менее могут быть неспособны иметь потомство друг с другом. Таким образом, являясь одним фенетическим видом, они принадлежат разным биологическим видам. Примером является фруктовая мушка Drosophila с ее двумя (неспособными к скрещиванию) категориями, D. pseudoobscura и D. persimilis. Эти два организма являются практически неразличимыми фенетически, поэтому они образуют один фенетический вид, но, будучи нескрещивающимися, составляют два биологических вида. Существуют и другие определения того, что значит быть видом, и применение критериев, которые в них предлагаются, мутит воду еще больше. Экологическая концепция видов принимает во внимание важность роли окружающей среды, ресурсов и опасностей, из нее исходящих. Она определяет вид как группу организмов, использующую одиночную экологическую нишу. Опознавательная концепция видов принимает во внимание способность организма опознавать потенциального партнера для спаривания. Преимущество этого определения, тесно связанного с биологической концепцией видов, состоит в том, что если возможность спаривания зачастую приходится лишь предполагать, то опознавание часто можно наблюдать непосредственно. Возможны случаи появления нового вида, когда одна группа организмов не может более опознавать своих недавних супругов в качестве потенциальных партнеров. Опознавание не обязательно происходит по внешнему виду: растения и животные сообщаются между собой различными путями, включая звук, а также более сдержанный или даже, по нашим понятиям, бессознательный путь испускания и детекции химических веществ, которые мы называем феромонами и которые человеческие существа иногда включают, по абсолютно тем же мотивам, в свои духи и лосьоны. И наконец (наконец — лишь в этом кратком обзоре, поскольку имеются и другие определения), существует филогенетическая концепция видов, в которой вид определяется как группа организмов, имеющая общего предка, и отличающаяся от других групп по крайней мере одним признаком. В соответствии с этим определением, члены двух разных филогенетических видов могут различаться всего одним признаком и быть способными иметь общее потомство. Нет сомнения, что виды эволюционировали и продолжают эволюционировать. Свидетельством эволюции в прошлом являются ископаемые останки, которые разворачивают во времени замечательную последовательность образов населявшей Землю жизни. Эта картина неполна, ведь сегодня ни один музей — а музеи обычно гораздо больше пекутся о своей собственности, чем сыра земля — не имеет образцов каждого исчезнувшего вида. Но она достаточно полна для того, чтобы мы могли проследить сквозь время происхождение живых существ, включая наше собственное возникновение в — сюда не подходит клише «далекое и туманное» — недавнем и ярком прошлом. Наука об ископаемых останках и их интерпретации в контексте истории жизни на Земле носит название палеонтология. (Английское слово «fossil» — ископаемое, произошло от латинского слова «fodere», выкапывать, через слово «fossile», означающее «выкопанный».) Первые охотники за ископаемыми, придерживаясь точки зрения платоников, считали, что ископаемые являются образом идеальной формы, созданной действием некоего рода формотворящей силы — vis plastica. Однако теперь мы знаем, что ископаемые состоят из минеральных частей скелетов (кости содержат главным образом фосфат кальция вкупе с белковым хрящем) и зубов (тоже фосфат кальция с разными видами твердых покрытий). Ископаемые находят в осадочных породах, породах, сформированных путем осаждения и спрессовывания минералов, как, например, известняк. Изверженные породы, породы, которые вытекли на поверхность с больших глубин, никогда не населены ископаемыми. Некоторые ископаемые находят в метаморфических породах, которые представляют собой осадочные или изверженные породы, видоизмененные под действием высоких температур и давлений. Некоторые ископаемые являются органическими веществами, такими как древесина, которая минерализуется, когда вода просачивается в нее и заполняет внутренние полости окаменевшими отложениями. Органический прародитель может совсем исчезнуть к тому времени, как мы наткнемся на него, и то, что мы откопали, окажется трехмерной минеральной копией оригинала. Раковины часто сохраняются, но карбонат кальция в форме арагонита, из которого они состояли, превращается в более твердую и плотную форму, известную как кальцит. Органические вещества не сохраняются таким способом, но отпечатки перьев (белок жесткого типа) и мясистых частей (состоящих из белков мягких видов, смазанных жирами) часто обнаруживают в породе, в которую было вмуровано ископаемое. Некоторые крошечные существа хранятся неповрежденными в затвердевшей смоле, которую мы называем янтарем. Более крупные создания, например мамонтов, находят в сохранности вмороженными в ледники. Земля под нашими подошвами живет, в ней происходит безостановочный подъем вещества из расплавленных областей внизу, создающий новые участки литосферы, внешнего твердого футляра расплавленной внутри Земли. Восходящие струи магмы заставляют литосферу расширяться в зоне ее изливания и затем в отдалении снова погружаться вниз в желоб субдукции. В этот конвейерный ремень включены коровьи лепешки коры, называемые континентами, которые перемещаются таким путем по поверхности глобуса. Это процессы тектоники плит впервые предложены высокомерному миру немецким геологом Альфредом Вегенером (1880-1930) и обоснованны в его труде Происхождение континентов и океанов (1915), но лишь около 1960 г. приняты к разработке, которая показала, что считавшееся доселе неподвижным жесткое дно океана может раздвигаться, преобразуя внешний вид Земли (рис. 1.3). Они вызывают также локальное прогибание континентальной коры, имеющее последствия широкого диапазона, от орогенеза (горообразования) до формирования каньонов, предгорий и долин.
Не вызывает удивления, что среди этого великого переселения в ряде случаев происходит перемешивание геологических слоев, и то тут, то там ископаемые одного поколения попадают под ископаемые другого, а более древние ископаемые организмы вымываются откуда-нибудь, чтобы перемешаться со своими потомками. Обычно мы можем выявить эти очевидные несоответствия, прослеживая форму слоя и наблюдая характер его прогибания. Однако, если мы примем во внимание мощь геологических событий в совокупности с буйными вмешательствами климата, когда океан замерзает на века, ледники скребут взад-вперед, а затем, когда лед отступает, огромные стометровой высоты цунами снова наполняют океан талой водой, можно лишь удивляться, что сохранились вообще хоть какие-то следы нашего далекого прошлого. Глобальная война — Земля против организма, организм против организма — свирепствует над останками жизни, и найти хотя бы один зуб — уже удача. Но мы находим гораздо больше, чем один зуб. Удача в смерти, если такая вещь существует, позволила динозаврам умереть, когда не стало чем пообедать, позволила им погрузиться в ил, завернуться в осадочные породы и в нужное время быть вытолкнутыми к дневному свету, когда эрозия удалила покровы. Богатейшие ископаемые останки состоят из морских беспозвоночных с твердыми скелетами, обитавших на мелководье. Хуже всего представлены организмы без скелетов и легко повреждаемые создания, такие как птицы. Некоторые ископаемые встречаются в огромных количествах: меловые холмы сложены из ископаемых останков одноклеточных водорослей, известных как кокколитофориды (рис. 1.4). Эти ископаемые отлагаются и сегодня, около 1,4 миллиарда килограммов кокколитофорид оседает ежегодно. Их присутствие в морской воде отчасти отвечает за ее прозрачность. Действительно, летом 1997 и 1998 гг. все Берингово море меняло цвет от глубоко синего к аквамариновому, ибо оно цвело миллионами кокколитофорид в их краткий, но безмолвный момент наслаждения жизнью по пути к превращению в будущие возвышенности.
Ископаемые записи, даже будучи вызывающе неполными, в высшей степени убедительно свидетельствуют об эволюции, в ходе которой виды приходят и уходят, одни виды порождают другие, а иные вымирают, и это все подобно кусту с ветками, которые ветвятся, побегами, которые отмирают, и с листьями, создающими подобие современной биосферы. Такие записи будто предназначены для того, чтобы показать похожую на куст историю биосферы с правдоподобными, хотя и допускающими иногда разные толкования, линиями наследования. Однако существуют альтернативные объяснения ископаемых данных, и, поскольку предмет столь важен для понимания нашего места в природе, мы должны исследовать его. Одной из альтернатив эволюционизму является трансформизм; мы займемся этой гипотезой позднее. Здесь мы рассмотрим другую альтернативу, креационизм, в которой каждый вид остается навсегда неизменным, если не считать незначительных вариаций. Согласно креационизму вид, уже готовый, разумно спроектированный, с дышащей в нем жизнью, вызван к существованию несотворенным, всемогущим, достойным поклонения Творцом. Возможно, вид будет вечным, а может быть, вымрет, уступая помещение для нового проявления непостижимой прихоти Творца. Творец имеет неограниченные возможности проектировать и создавать животных, а также, очевидно, неистощим в вариациях на тему их способностей мучить, калечить и убивать друг друга. Среди этих клыков, сочащихся ядом, конечно, находится и Человек, образ Творца, обладающий свободой воли. Креационизм, включая его прозрачно закамуфлированную разновидность «Разумный План», не является наукой: это непроверяемое утверждение, инспирированное антинаучной и религиозно мотивированной повесткой дня. При максимальной снисходительности креационизм может выполнять роль, аналогичную роли галилеевского Простака, роль литературного приема для демонстрации того, что научный подход, в данном случае эволюционизм, наилучшим образом объясняет предмет. Важно, что от науки постоянно требуют приводить объяснения; проблема же креационизма в том, что его защитники не осознают, что являются не более чем Простаками. Их бесконечная и раздражительная докучливость, даже хуже, искажение очевидного являются бесплодными и надоедливыми и создают для молодых опасность того, что их глаза станут слепыми для восприятия подлинной славы творения. Итак, каковы аргументы против креационизма? Их так много, что изложение всех переполнило бы эту главу. Я дам почувствовать их аромат, приведя только три из них. Первый: многочисленные новые виды появились в последнее время, и это настоятельно внушает нам мысль, что так же происходило и прежде, что ископаемые не являются просто остаточными костями, свидетельствующими скорее о вымирании, чем об эволюции. Второй: не раз заявлялось, что эволюция не имеет предсказательной силы, поэтому не может быть проверена и, следовательно, не в большей степени является разновидностью науки, чем креационизм. Это утверждение неверно. Тот факт, что эволюция происходила, появился на свет благодаря наблюдениям останков вымерших разновидностей макроскопических животных. В двадцатом веке стало очевидным, что эволюция может быть прослежена и на молекулярном уровне. Эффективным оказывается предсказание того, что детали молекулярной эволюции должны соответствовать деталям макроскопической эволюции. Выясняется, что так оно и есть, не существует ни одного примера, в котором следы молекулярных изменений расходились бы с данными наших наблюдений целостных организмов. Третий: одна из узаконенных проверок соблюдения авторских прав состоит в том, чтобы установить, воспроизводит ли продукт погрешности, внесенные, иногда преднамеренно, в образец, который он копирует. Картографы иногда вносят небольшие ошибки — например, лишний домик в ландшафте, — чтобы подловить плагиаторов. В биологии имеются два вида ошибок, подверженных плагиату. В одном — эволюция начинается в глупом направлении (которое не было предусмотрено) и далее вынуждена нести последствия этого. Часто цитируемым примером является глаз млекопитающего, поскольку в своей эволюции он оказался пойманным в придурковатую конструкцию, которую мог бы придумать Придурковатый Конструктор, с кровеносными сосудами, лежащими перед ретиной. Сосуды вынуждены поэтому покидать глаз, продираясь через ретину и создавая слепое пятно. Природа следует этой конструкции и в настоящее время. Другой вид ошибки возникает на молекулярном уровне, в форме, например, псевдогенов, которые воспроизводят нефункциональные полоски мутировавшей ДНК, эквивалент ложного домика на картах. Но вернемся к науке и установленному факту эволюции. Микроэволюция — это процесс развития мельчайших модификаций. Макроэволюция — это порождение новых видов и таксонов более высокого ранга (отрядов, семейств и т.д.) в результате накопления изменений, вносимых микроэволюцией, это процесс, называемый филетическим градуализмом. Как мы уже отмечали, экспериментальные свидетельства такой постепенной эволюции затуманены предполагаемой неполнотой ископаемых записей, в которых часто отсутствуют переходные формы, которые следовало бы ожидать. Существуют два возможных объяснения этому. Одно состоит в том, что переходные формы существовали, но исчезли без следа. Альтернативное объяснение заключается в том, что филетический градуализм некорректен, что ископаемые записи более полны, чем мы думали, и что видообразование (порождение новых видов) происходит вспышками в течение нескольких тысяч лет, следующих за долгим периодом покоя или «стазиса». В высшей степени спорная теория прерываемого равновесия была предложена Нильсом Элдриджем и Стивеном Гулдом (1941-2002) в 1972 г. В этой теории предполагается, что малое изолированное сообщество испытывает взрыв модификации в процессе аллопатрического видообразования («аллопатрическое» означает просто, что изменение происходит в географическом районе, отличном от места обитания предков). Поэтому место обитания предков вряд ли содержит образцы промежуточных форм, и ископаемые останки нового вида могут быть найдены там лишь в том случае, если полностью развившийся новый вид, расселяясь, попал туда вновь: понятное отсутствие промежуточных форм усиливает впечатление внезапности перехода между двумя формами. Филетический градуализм и прерываемое равновесие, в том виде, в котором они были предложены первоначально, вероятно, лучше всего рассматривать как противоположные концы спектра возможностей. Их не представляется уместным считать конкурирующими моделями эволюции, скорее это метки на шкале прибора, измеряющего скорость, с которой происходит видообразование. Некоторые события, появление некоторых видов, соответствуют показанию стрелки, близкому к градуализму, а для иных, для появления других видов, стрелка склоняется к прерыванию. В высшей степени трудно характеризовать скорость эволюции видов и быть уверенными, что ископаемые данные являются полными. Нельзя сказать, что более современные версии прерываемого равновесия не являются спорными, поскольку они разрабатывались за пределами простого контрапункта «быстро-медленно» ранних модификаций; в частности, были предложены механизмы поддержания стазиса и переключения к эпизодам быстрых изменений. Философская установка теории тоже спорна: в то время как дарвинизм предполагает, что видообразование есть накопление изменений в результате адаптации, прерываемое равновесие рассматривает видообразование как движущую силу адаптации. Наличие таких споров не стоит интерпретировать как ошибочность теории естественного отбора (и тем более факта эволюции): они лишь являются признаком жарких дебатов вокруг деталей одного из наиболее важных процессов в мире. Далее необходимо подчеркнуть одно обстоятельство. Эволюция не обязательно ведет ко все большей изощренности: направление эволюции не всегда идет вверх. Организм может обнаружить, что способен ускорить свою репродуктивную активность и тем самым более успешно заселить Землю, если он отбросит большую часть своего социального или анатомического багажа. К чему беспокоиться и изобретать множество способов социальной активности, если можно выполнить главную работу воспроизводства без них? Более того, среда обитания может измениться, и остаточные особи непреуспевшего вида могут вдруг обнаружить, что пришел их час, что в изменившихся условиях они могут по плодовитости обставить своих до сих пор более успешных конкурентов. Оболочник, морской сифон (Ciona intestinalis), реализует другое решение, становясь в конце концов подобием картофельного клубня. Эта маленькая пасть является в своей личиночной фазе подвижным охотником и поэтому нуждается в мозге. Однако, когда он находит подходящую нишу, в которой может себя закрепить, чтобы стать сидячим, ему больше нет нужды думать, поэтому он поедает свой собственный энергетически обременительный мозг. Мозги являются великими потребителями энергии, и это неплохая идея — избавиться от своего мозга, когда вы обнаруживаете, что он вам больше ни к чему. Откуда берется все это богатое многообразие живых организмов? Уильям Пэйли, как мы видели, не имел сомнений на этот счет, поскольку был уверен, что каждый вид является творением Бога, и это все. Жан Батист Пьер Антуан де Моне, шевалье ле Ламарк (1744-1829) тоже думал, что знает, но его интеллект был настолько более замечательным, чем интеллект Пэйли, что он бился над проблемой обнаружения механизмов. Ламарк, сначала солдат, затем банковский служащий, потом ботаник-ассистент и, наконец, профессор насекомых и червей, провел жизнь в бедности, а последние несколько лет и совершенно слепым. Бедность преследовала его даже в смерти, он был похоронен в арендованной могиле, чтобы быть выкопанным, когда срок временного владения через пять лет истек, и совершать путь к новому захоронению, в результате чего останки его рассеялись. Его имя сегодня больше ассоциируется с презрением, чем с уважением, хотя он заслуживает уважения как основатель биологии беспозвоночных (термин, который придумал он) и за то, что он по крайней мере пытался отыскать объяснение существования видов. Он начал публиковать свои умозрения — это не было чем-то большим и, конечно, не было научной теорией — о механизмах эволюции в 1801 г., но его наиболее полный отчет был представлен в труде Philosophic zoologique (1809). Ламарк считал, что все организмы вовлечены в метафизический поиск пути к совершенству и проходят путь трансформаций из одноклеточного зародыша, содержащего некоего рода платоновскую сущность вида. Этот поиск направляют «нервные флюиды» различных трудно определимых разновидностей, которые питают органы, делающие зарядку, и морят голодом органы, уклоняющихся от нее. Он также умозаключил — и это та самая идея, за которую его больше всего помнят, хотя он сам, вероятно, считал ее лишь малой составляющей своих полных тезисов — что однажды приобретенные свойства наследуются. Хорошо известен его пример удлинения шеи жирафа, когда тот пытался достать до более высоких листьев и стать еще более совершенным жирафом, причем достигнутое одним поколением удлинение наследовалось благодарными потомками в следующем колене. Можно осмеивать примитивную наивность данной идеи, однако, пока молекулярная биология не исключила возможность любого механизма такого наследования, эту концепцию было трудно опровергнуть. Ламаркианские взгляды, которые скорее относятся к трансформизму, чем к эволюционизму, упорно продолжали существовать и в двадцатом веке. Шуточное их опровержение является общепринятым, но непригодным: то, что обрезание крайней плоти у многих поколений евреев не привело к ее атрофии, не является аргументом, поскольку маленькие мальчики не старались потерять крайнюю плоть. В знаменитой серии неприятных экспериментов влиятельный немецкий биолог Август Вейсман (1833-1914) отрезал хвосты многим последовательным поколениям мышей и не обнаружил никакого уменьшения длины хвоста у последующих поколений. Все такие калечащие эксперименты — а их было много, как случайных, так и преднамеренных, — хотя и подходили для опровержения того, что приобретенные свойства наследуются, не соответствовали центральному тезису трансформизма, точке зрения Ламарка, утверждавшей, что главным является стремление, ибо лишь тогда трансформирующие сущности вступают в дело. Роберт Чемберс (1802-71) в своей книге The vestiges of Creation, опубликованной в 1844 г., уловил проблеск возможного объяснения. Он осознал важность мутаций, но доказывал, что новые виды причудливым образом возникают из случайно появившихся уродливых птиц. Так, если бы рыба непостижимым образом родилась с крыльями, перьями и клювом, то биосфера приобрела бы нечто, напоминающее обитающих в ней птиц. Примерно в это же время по воле и на средства преподобного Генри Эгертона, восьмого и последнего графа Бриджуотера, были изданы Bridgewater treatises (Бриджуотерские трактаты), сборник работ, созданный для того, «чтобы показать могущество, мудрость и милость Бога, проявленные в его Творении, иллюстрируя этот труд всеми разумными доводами, как, например, разнообразием и строением Божиих тварей». Эта книга явилась средством для выражения многих идей того времени. Среди включенных в нее статей были «Адаптация внешней природы к моральному и интеллектуальному устройству человека» Томаса Чалмерса (1833) и «Адаптация внешней природы к физическому состоянию человека» Джона Кидда (1837). С современной точки зрения содержание обеих статей в точности противоположно тому, что мы считаем верным сегодня. И вот, в последней части текущей главы, герой эволюции Чарлз Дарвин (1809-82) наконец нерешительно вступает на эту страницу. Успех Дарвина в решении вопроса о происхождении различных видов организмов можно проследить с момента его погружения в мир природы на период с 1831 г. по 1836 г., когда он номинально служил компаньоном, а фактически натуралистом на корабле Ее Величества «Бигль», при капитане Роберте Фицрое, незаконнорожденном потомке короля Карла II. Фицрою был нужен джентльмен в качестве компаньона в долгом одиноком плавании не в последнюю очередь для того, чтобы избежать судьбы своего предшественника на корабле, пустившего себе пулю в лоб. Страшился он того, что могло оказаться и наследственной склонностью, так как несколько лет назад его дядя, министр внутренних дел виконт Кэслро, в припадке депрессии перерезал себе горло. Погружение в избыток несметных, по-видимому, данных часто бывает прелюдией к зачатию открытия, к подсознательной неотвязной работе где-то в скрытых областях мозга, к поиску моделей и, наконец, к заключительному прорыву осознанной мысли, порождающему самое драгоценное из личных переживаний ученого, восклицание «эврика». За пять лет своего путешествия Дарвин провел много месяцев на суше, обычно желанной ему как отдых от морской болезни, редко покидавшей его на борту утлого суденышка.[2] Самым знаменитым из его временных пристанищ в течение пяти недель с 15 сентября 1835 г. были Галапогосские острова («острова черепах»), у побережья Эквадора в Тихом океане, куда «Бигль», как и столь многие до него, зашел для сбора огромных местных черепах ради их свежего мяса, необходимого для совершения следующего морского перехода. С тех пор на более крупных островах охота на черепах привела к полному их исчезновению, и лишь несколько особей выжили на островах небольших. Галапагосы являются цепью вулканических островов, которую в другое время другой посетитель, Герман Мелвилл, с гораздо меньшей проницательностью, чем довелось проявить Дарвину, характеризовал как «мелкую распродажу куч пепла, наваленных здесь и там за чертой города». Но даже Дарвин не оценил значение этого посещения до тех пор, пока острова не оказались далеко за кормой, ибо записал, что «трудно вообразить тропические острова, столь бесполезные для человека». Обволакивающие туманы и переменчивые течения, окружавшие острова, прилепили к ним прозвище Los Encantadas («заколдованные»), какими, конечно, они и были из-за того, что метафорический туман, который до тех пор окутывал происхождение видов, начал рассеиваться, когда Дарвин, с аппетитом уплетая плоть черепах, которых он помогал разделывать, размышлял над различиями между трупами птиц, собранными им на разных островах (он посетил только Сан-Кристобаль, Флореану, Изабеллу и Сантьяго). И он смог записать: Несколько островов обладают своими собственными видами черепах, дроздов-пересмешников, вьюрков и многочисленных растений, эти виды имеют одни и те же общие среды обитания, занимая аналогичные территории, и, очевидно, заполняя то же место в экономике природы… Это меня поражает.Как мы уже отмечали, острова оказались решающими для фомулирования теории, которую Дарвин смог в надлежащее время назвать теорией естественного отбора. Острова не только упрощают экосистему и поэтому облегчают обнаружение различий, но также эффективно изолируют популяции, помогая таким путем развитию и адаптации их различных вариаций. Набитому черепахами и пораженному Дарвину пока не хватало искры, чтобы мысль его появилась на свет. Искра вспыхнула, как он утверждал впоследствии, 28 сентября 1838 г., когда он все еще осмыслял гору информации, которую скопил за свое долгое путешествие. Читая для развлечения Essay on the principle of population (Опыт о законе народонаселения) Мальтуса (1798), в котором элегантный и утонченный Реверенд Томас Мальтус (1766-1834), профессор политической экономии, нанятый обучать экономике служащих Ост-Индийской компании, доказывал, что человечество обречено, поскольку население растет быстрее, чем запасы пищи, следовательно, человечество неизбежно исчерпает свои ресурсы. Дарвин позднее вспоминал: Благодаря хорошей подготовке к тому, чтобы оценить борьбу за существование, которая обнаруживается повсюду при длительном наблюдении за поведением животных и растений, меня сразу осенило, что под действием этих обстоятельств благоприятные изменения имеют тенденцию к сохранению, а неблагоприятные — к разрушению.Томас Хаксли (1825-95), верный бульдог Дарвина, смог позднее сказать: «Было бы в высшей степени глупо не догадаться об этом». Почти добрых двадцать лет Дарвин размышлял над этими наблюдениями, постепенно выстраивая свою теорию естественного отбора, подбирая иллюстрации, так никогда до конца и не утратив веру в ламаркианское наследование приобретенных признаков и опасаясь последствий публикации. Он начал писать отчет о своих идеях в 1856 г., намереваясь, подобно доктору Кэсэбону, герою Джорджа Элиота, сделать его огромным и авторитетным. Но его планы были сорваны, поскольку существовали и другие посетители островов, читавшие Мальтуса. Дарвин пришел в смятение, получив от Альфреда Рассела Уоллеса (1823-1913) манускрипт, носящий название «О тенденции разновидностей к неограниченному отклонению от первоначального типа». Уоллес был далеким потомком героя Шотландии Уильяма Уоллеса и с 1848 по 1852 г. широко обследовал бассейн Амазонки в качестве профессионального собирателя образцов. После непродолжительного ожидания в Европе он решил возобновить свою все более прибыльную, но и более напряженную карьеру собирателя и, избрав Малайский архипелаг (Индонезийский архипелаг) в качестве места назначения, в 1854 г. прибыл в Сингапур. В феврале 1858 г., после нескольких лет путешествий и собирательства, подвергшись атаке малярии на Молукках (на каком острове, точно не известно, возможно это был либо Джилоло, либо Тернате), он понял — как Дарвин, — что идеи Мальтуса содержат ключ к объяснению эволюции. Дарвин очутился в затруднительном положении, это была именно та идея, которую он вынашивал два десятилетия, и приоритет, казалось, уплывал у него из рук. Он обратился за советом к своим друзьям, сэру Чарлзу Лайеллу и ботанику Джозефу Гукеру. Не имея возможности проконсультироваться с Уоллесом, они решили представить эссе последнего и сведенные вместе заметки Дарвина на следующем заседании Линнеевского общества в Лондоне 1 июля 1858 г. С этого момента естественный отбор перестал быть секретом. С этого момента Дарвин отказывается от своего выдающегося опуса, с яростью сокращает его запланированный объем, проявляя милосердие к читателю, и публикует в ноябре 1859 г. работу О происхождении видов, или, более точно, если уж полностью восстанавливать по-викториански готическую декорацию, О происхождении видов путем естественного отбора или сохранении благоприятствуемых пород в борьбе за жизнь. Даже сам Дарвин находил это название немного нескладным и в последующих изданиях (их было еще пять) он зашел уже так далеко, что выкинул предлог О. Дарвин замечал: Я имел в виду две отчетливых цели, во-первых, показать, что виды не были сотворены по отдельности, и, во-вторых, что естественный отбор являлся главным фактором изменения.Наше внимание должно теперь сфокусироваться на Дарвине, общепризнанном первооткрывателе естественного отбора. Но было бы неверно в этой связи полностью игнорировать Уоллеса, не в последнюю очередь за благородство, с которым он фактически уступил приоритет Дарвину. Однако позднее, в своей очень долгой жизни, Уоллес проявил некоторые свойства, уменьшившие его вес в этой области науки. Он так никогда и не принял мысль, что человек мог эволюционировать без малейшего божественного дуновения и руководства. Он желал ограничить естественный отбор эволюцией форм, не способных осознавать формирующее влияние некой высшей силы. И наконец, к вящему ужасу друзей, его ум окончательно потерялся на бесконечных и мрачных тропинках спиритуализма. Естественный отбор — это очень простая идея, однако она очень сложна в приложении, поскольку исследования, которые необходимо провести, требуют величайшей осторожности. Говоря вкратце, черепах на острове недостаточно; чтобы рассмотреть влияние естественного отбора на виды черепах, мы должны рассмотреть его влияние на всю ближайшую флору и фауну, а также физические условия и климат в зоне их обитания. Эволюция черепах будет иметь последствия для их конкурентов и хищников, которые, в свою очередь, будут воздействовать на черепах. В противоположность простым линейным системам, в которых влияние осуществляется посредством простой цепочки команд, биосфера чрезвычайно богата нелинейными системами, в которых изменения организма сами воздействует на него, поскольку его эволюция изменяет среду. Развитие во времени нелинейных систем очень трудно прогнозировать, и неудивительно, что эволюция не способна предсказать будущее биосферы, представляющей собой апофеоз нелинейной сложности. Здесь я очерчу облик некоторых идей, характеризующих современный синтез, или неодарвинизм, которые показывают, как случилось, что в начале двадцатого века идеи генетики, заложили фундамент для идей, связанных с наблюдениями естественной истории. Теория естественного отбора не была вполне признана вплоть до 1930-х гг., когда современный синтез утвердился. Как уже было сказано, в этой главе я ограничиваюсь главным образом феноменологией, откладывая молекулярный базис эволюции до следующей. Естественный отбор опирается на три принципа: 1. Существуют наследуемые генетические изменения. То есть особи данного вида не являются идентичными клонами; внутри вида имеется генетический шум. У Дарвина не было концепции механизма наследования, и он оказывал предпочтение теории перемешивания, в которой характеристики спаривающихся родителей попадают в нечто вроде миксера. Эта неосведомленность об истинных механизмах и склонность к механизмам, которые, как быстро установили критики, не могли обеспечить эволюцию, и были главным препятствием для признания его идей. История могла бы быть другой, если бы Дарвин дал себе труд распечатать и прочесть письмо ничем не прославленного монаха, Грегора Менделя, который фактически вручал ему ключ. 2. Родители производят избыточное потомство. То есть, как бы следуя Мальтусу, родители производят потомков больше, чем способно выжить. Некоторые виды, такие как слоны, рождают только одного отпрыска, и он может умереть, другие, такие как лягушки, производят тысячи, из которых выживает, возможно, только один. Избыточное производство потомства случается реже у больших, сложных организмов, способных позволить себе годы родительской заботы, таких как слоны или, может быть, принадлежащие к среднему классу родители из стран Востока. 3. Успешные потомки лучше адаптированы к окружающей среде. «Успех» это больше чем простое выживание; это еще и способность к продолжению рода. Этот принцип является пружиной злосчастной и неверно понятой фразы правого либерала девятнадцатого века Герберта Спенсера «выживание наиболее приспособленных», которую он бросил (примерно в 1862 г.) в связи с развиваемой им теорией социального дарвинизма, где необдуманно распространил утонченные идеи естественного отбора на динамику общества и открыл дверь евгенике, исключению практически всех форм государственного вмешательства и расизму. Как все удачные лозунги, «выживание наиболее приспособленных» запоминается, и Дарвин испытывал соблазн употребить его в более поздних изданиях, но это снижало ценность изящной идеи, лежащей в основании его теории. Когда мы рассматриваем естественный отбор, необходимо иметь в виду, что он абсолютно локален и ограничен во времени. Естественный отбор полностью погружен в настоящее и совершенно лишен предвидения. Если бы сегодняшняя адаптация принимала жалобы из будущего, это была бы плохая услуга будущему: естественный отбор не может заранее знать, что он ведет вид в эволюционный тупик; он не может, конечно, и вообще ничего знать заранее, даже на один день вперед. Естественный отбор живет моментом, он в высшей степени гедонистичен. Мы уже упоминали в качестве примера глаз млекопитающих: по капризу эволюции первоначальное светочувствительное пятно, которое в процессе развития сумело стать главным органом восприятия, в начале развития имело кровеносные сосуды на своей передней стороне, что в результате привело к перекрыванию ими ретины (рис. 1.5). Светочувствительность — такое мощное оружие для преследования и убегания, что более ценным для организма оказалось продолжить существование с этим несовершенным устройством, чем отказаться от преимущества и опрокинуть заведенный порядок ради ожидания лучшего глаза через миллион лет. Глаз головоногих более удачен в этом отношении (но не в других), поскольку развивался по эволюционному пути, на котором кровеносные сосуды оказались лежащими за светочувствительной ретиной.
Другим примером является неудобная конструкция трубок внутри нашего горла, где дыхательный и пищевой проходы пересекаются, что открывает возможность заклинивания. Проходы пересекаются потому, что у нашего древнего предка, рогозуба, рыбы, имеющей легкие, отверстие, используемое для дыхания у поверхности было очень удобно расположено в верхней части головы и вело в общее пространство, деля его с пищевым проходом (рис. 1.6). Это устройство не подверглось изменению, несмотря на его опасность. Очевидно негигиеничная экономия, приведшая к использованию пениса как для копуляции (включая, особенно у людей, сопутствующие ритуалы), так и для мочеиспускания, имеет похожую эволюционную причину, более того, трубка, ведущая от яичек к пенису, делает петлю с неудачной стороны от трубки, соединяющей почки с мочевым пузырем.
Естественный отбор по существу непредсказуем, так как он является результатом подчас конкурирующих тенденций, и способы адаптации, на первый взгляд предпочтительные, оказываются недостижимыми. Самым малым примером является аппендикс человека. Для нас аппендикс представляет опасность, поскольку он может воспалиться и привести к смерти. Аппендицит возникает, когда инфекция вызывает опухание, которое сдавливает артерию, снабжающую аппендикс кровью. Стабильный поток крови через аппендикс защищает его от роста числа бактерий, так что любое уменьшение потока помогает инфекции, что приводит к еще большему сдавливанию. Если поток прекращается совсем, бактерии благоденствуют, а аппендикс лопается. Маленький аппендикс чаще подвержен этой цепочке событий, чем большой аппендикс, поэтому аппендиксы подвергаются селективному давлению, которое становится на сторону большого аппендикса в том смысле, что опаснее начать уменьшать его, чем оставаться с тем, что у нас есть. Следовательно, несмотря на опасность, эволюции крайне трудно удалить аппендикс. Естественный отбор является гонкой вооружений. Гипотеза Красной Королевы — это идея о том, что хищники и жертвы вовлечены в постоянное сражение, в котором хищники вырабатывают все более совершенные стратегии и техники преследования, а жертвы поступают подобным же образом. (Красная Королева давала Алисе инструкцию продолжать бежать еще быстрее, чтобы оставаться на месте.) Более острые зубы повсюду приводят к появлению более толстых шкур или быстрых ног, что в свою очередь поощряет появление еще более острых зубов. Естественный отбор является также и зеркалом окружающей среды. Выразительный пример воздействия физического окружения на направление естественного отбора дает независимое возникновение сходным образом адаптированных организмов в удаленных друг от друга частях мира. Ничто не делает этот процесс конвергентной эволюции более впечатляющим, чем возникновение сумчатых версий плацентарных млекопитающих; в древние времена эмбрион развивался в основном во внешней полости, а в более поздние он развивается главным образом в сумке (in utero). Сумчатые версии появились, когда Австралия откололась от Антарктики в цейнозойскую эру, около 65 миллионов лет назад, и поплыла на Север, как Ноев ковчег, со своей изолированной экосистемой. Североамериканский волк (Canis lupus), плацентарное млекопитающее, подобен по внешнему виду сумчатому тасманийскому волку (Thylanicus cynocephalus). Естественный отбор, осваивая подходящие ниши, привел ко многим аналогичным решениям (рис. 1.8): млекопитающий оцелот (Felispartialis) напоминает сумчатого тигрового кота (Dasyurus maculatus), белка летяга (Glaucomus volans) передразнивает сумчатую летягу (Petaurus breviseps), североамериканский лесной сурок (Marmota monax) является тенью вомбата (Vombatus ursinus platurrhinus), а обычный крот (Scalopus aquaticus) выглядит как сумчатый крот (Notoryctes tryphlops). Даже у домашней мыши (Mus musculus) есть свой сумчатый призрак-двойник в лице желтоногой мыши (Antechinus flavipes).
Единственным моментом, позволяющим осмыслить все эти взаимосвязи, является появление примерно 3,5 миллиона лет назад вулканического Панамского перешейка между материками Северной и Южной Америк, которые раньше были отдельными фрагментами соответственно Лавразии и Гондваны. Это событие привело не только к межвидовым битвам, когда популяции млекопитающих Севера хлынули на юг и сражались за выживание с преобладающими популяциями сумчатых Юга, но перешеек нарушил также циркуляцию океанов и положил начало Ледниковому периоду, изменившему флору и фауну всей планеты. Однако эволюционная война это лишь одна из компонент движущей силы изменений, так как изменения физической среды также играют центральную роль в управлении эволюцией. Эти изменения включают освобождение ниш при массовом вымирании, что позволяет развиваться новым пузырькам популяций. Катастрофизм, идея о том, что мир подвержен внезапным подвижкам, драматически представленная в мифе о Великом потопе, пришелся по вкусу, как причина изменений, высоко интеллектуальному французскому анатому и основателю палеонтологии позвоночных, барону Жоржу Леопольду Кретьену Фредерику Дагоберу Кювье (1769-1832), чьи имена многочисленны как геологические эры, но вышел из моды, когда геология закончила период становления. Начало рационализации геологии своими усилиями положил Джеймс Хаттон (1726-97) в труде Теория Земли (1795) и энергично продолжил сэр Чарлз Лайелл (1797-1875) в его трехтомных Принципах геологии (1830-33; Дарвин вез с собой на «Бигле» их экземпляр). Хаттон и Лайелл предпочитали униформитаризм, который, основываясь на большом числе свидетельств, полученных из анализа слоев, считал, что физическая природа Земли подвергалась медленной и постоянной трансформации. Однако теперь мы знаем, что катастрофы, конечно, были. Наиболее известной является столкновение с астероидом, которое почти полностью уничтожило жизнеспособных, но генетически недостаточно гибких динозавров. Эти огромные твари были обречены, когда искусственная ночь, произведенная пылью, окутавшей планету, привела к недостатку растительной пищи, или, возможно, они были сожжены миром, в котором концентрация кислорода в атмосфере была значительно выше, чем сейчас. Их кончина освободила мир для волны млекопитающих. Нам придется далее делать ссылки на некоторые геологические эры и периоды, на которые была разделена история нашей пластичной планеты (рис. 1.9). Имена, присвоенные им, довольно прихотливы, но Уэльс и запад Англии представлены в них очень неплохо: Кембрия (для Кембрийского периода) — древнее название Уэльса, Ордовики и Силуры (для Ордовикского и Силурийского) — имена доримских племен Уэльса, и Девон (для Девонского). Названия эпох, на которые разделены некоторые геологические периоды, имеют несколько расхлябанный вид: они включают Палеоцен («старый современный»), Эоцен («рассвет современного») и Олигоцен («мало современный»). Добавлю в скобках, что этимология других имен, по мере их возникновения, окончательно уничтожила остатки ранних попыток придать названиям периодов систематичный характер, как, например, Триасовый, Третичный и Четвертичный периоды.
Роковое вымирание динозавров в конце Мелового периода — только наиболее известное из по крайней мере пяти главных событий. Среди когда-либо произошедших катастрофических событий находится то, которое привело к концу Пермский период (Пермь, в восточной части европейской России), с исчезновением более 95 процентов видов морских животных. Ордовикский период был резко прерван 440 миллионов лет назад, Девонский — 350 миллионов лет назад, Пермский период — 250 миллионов лет назад, Триас — 205 миллионов лет назад и Меловой — 65 миллионов лет назад. Причины большинства этих вымираний все еще по большей части неизвестны, но недостатка в идеях нет, включая столкновения с астероидами и большие падения уровня моря, сопровождавшиеся глобальным похолоданием. Сопутствующие вымирания являются травматическими, но жизнь не унывает, и многообразие видов возвращается очень быстро: через 5-10 миллионов лет многообразие достигает уровня, имевшегося до вымирания, часто даже превосходя его. События вымирания сметают конкурентов, освобождают ниши, созревшие для колонизации, и поздравляют всех (кроме вымерших) с появлением благоприятных возможностей. Однако, хотя вымирания играют важную роль, не следует ее преувеличивать. Типичный для животных вид продолжает существовать около 2 миллионов лет, а вспышки вымирания случаются обычно каждые 20-30 миллионов лет, так что большая часть видов не подвергается вымиранию от катастроф. Злосчастная судьба динозавров таилась в том, что они оказались слишком живучими: они жили достаточно долго и дождались. В настоящее время мы, по-видимому, находимся в сердцевине массового вымирания нового типа, когда активность человека делает биосферу неблагоприятной для многих видов флоры и фауны, с которыми он делит ее, а возможно, и для него самого. Самоиндуцируемое вымирание этого типа может быть неотвратимым спутником «прогресса», так как с ультрапессимистической неомальтузианской точки зрения можно считать, что способность к самоуничтожению неизбежно обгоняет развитие интеллекта. Наиболее мрачный взгляд заключается в том, что общества еще могут выживать, если индивид способен убить одним ударом всего несколько тысяч человек (как это было до сих пор в человеческой истории), но ни одно общество выжить не может, когда технология развита до такой степени, что одна персона обладает мощью, достаточной для убийства десятков миллионов. Человеческое общество, возможно, как раз подошло к этой точке. Если это является общим правилом для населения всех планет, то у нас очень мало надежды, так художественно внушаемой оптимистичной научной фантастикой, надежды на исполнение космических чаяний человечества. Зато наше вымирание, по крайней мере, создаст благоприятные возможности для тараканов. Остается несколько центральных вопросов, касающихся всей этой богатой взаимосвязи между географией и генами, этой танцплощадки естественного отбора. Один из них — о природе объекта, на который воздействует естественный отбор. Воздействует ли он на ген, на индивида или на вид? Мы можем исключить вид как единицу отбора. Организмы ничего не делают ради своего вида. Так же как естественный отбор не видит будущего, он не видит и сообществ. Индивид конкурирует с другим индивидом и стремится к своему собственному успеху, не заботясь о пользе совокупности организмов, составляющей вид. Репродуктивная энергия индивида питает эгоистическое поведение и не имеет представления об альтруизме, бессознательном поведении, приводящем к самопожертвованию ради других. Нельзя отрицать, что многие виды поведения выглядят альтруистическими, и лишь когда мы более тщательно исследуем их, обнаруживается, что это волки в овечьих шкурах, и альтруизм на деле оборачивается красными каплями эгоизма на зубах и клыках. При взаимном альтруизме, представляющем собой управляющий идеальным человеческим обществом вариант социального контракта, организм получает прощение за свой эгоизм в обмен на сотрудничество с другими организмами в значительной мере для того, чтобы в тяжелые времена помогающий мог бы сам получить помощь. На более глубоком уровне следует понимать, что представители вида имеют общие гены, и помогая очевидным конкурентам по размножению, организм скрыто помогает распространению своих собственных генов. Этот тип альтруизма назван родственным отбором. Таким способом биолог-теоретик Дж.Б.С. Холдейн (1892-1964) выразил точку зрения, согласно которой он был бы рад утонуть, если бы это спасло двух его родных или десять двоюродных братьев. Каждый из его родных братьев разделял бы с ним половину его генов; его двоюродные братья разделяли бы одну восьмую (для компенсации хватило бы спасения восьми двоюродных братьев, спасение десяти дает уже генное преимущество). Контроль наших генов над нашим поведением предполагает, что мы можем смотреть глубже, чем уровень вида, глубже чем уровень индивида, прямо в глубь генов. Проблема этого взгляда состоит в том, что он редко находится во взаимно однозначном соответствии с поведением. Существует не только заговор сложности биосферы, но также и воплощение генотипа (генетической конструкции организма) в фенотипе (физических характеристиках организма). Некоторые организмы отказывают себе в радостях воспроизводства, но все же делают свой вклад в будущее, помогая вместо себя размножаться своим близким родственникам. Гены их матки (например, пчелиной) так близки к их собственным, что, помогая ей размножаться вместо себя, они достигают распространения и своих генов: ведь она может рассеивать копии их генов, освобождая их от необходимости самим беспокоиться об этом. Другой проблемой является отслеживание последствий конкуренции на одном уровне (например, индивидуальном) для другого уровня (видового). Может случиться, что выгода индивида вредоносна для группы. Поскольку индивид не обладает эволюционным предвидением, он может пренебрегать последствиями своего поведения для группы. Когда пищи недостаточно, некоторые индивиды продолжают размножаться и передают свои гены следующим поколениям; они не воздерживаются ради своего вида. В результате вид будет эволюционировать в направлении, задаваемом генным потоком эгоистичных копий. Современная эволюционная биология смотрит неодобрительно на групповой отбор, отбор на уровне вида или сравнимой группы индивидов: естественный отбор происходит на более низком уровне, и все эволюционные тенденции, которые, как кажется, указывают на межвидовый отбор, обычно могут быть прослежены до последствий отбора на нижнем уровне. Фактически, если исключить особый случай родственного отбора, нет никаких отчетливых примеров адаптации, которая недвусмысленно шла бы на благо группе: в лозунге «на благо вида» нет никакого содержания. Проблема единицы отбора может быть выражена разными способами, так как отбор максимален на определенном уровне. На самом низком уровне бытия, на уровне атомов, не имеет значения, кто кого препарирует, ибо атомы переживают и убийцу, и нанесение увечий и резню. На много более высоком уровне, возьмем, к примеру, царство Животных, также не имеет значения, кто кого забивает, поскольку выживание царства не связано с его составом. Влияние на выживание становится гораздо более существенным, когда мы достигаем уровня индивидов и их генов, так как разница между убийцей и убитым теперь жизненно важна. Небольшой сдвиг к краю шкалы приводит нас к видам: смерть индивида определенно влияет на будущее вида, поскольку обычно лучше иметь столь много производителей, сколь это возможно, и ваше выживание является вкладом, при условии, что вы репродуктивно состоятельны. Класс млекопитающих также несколько более склонен к выживанию, когда обедающим является млекопитающее, а обедом — кто-то другой, но «собака съедает собаку» — и вообще, млекопитающее съедает млекопитающее факт, почти нейтральный. Сдвигаясь от индивидов к противоположному краю шкалы, мы сталкиваемся с их генами, которые одинаковы для индивида и для вида. Обладает ли обед чьими-то генами большей или меньшей важностью, чем обед просто кем-то? Одним из подходов к выявлению единицы отбора является установление сущности, которая является потенциально бессмертной. Бессмертными являются атомы, но они представляют царство минералов, а не царство организмов. Компоненты, из которых формируется двойная спираль ДНК («нуклеотидные основания», которые мы обсуждаем в главе 2), не несут жизни внутри, так же как буквы алфавита не являются литературой. Даже если бы эти компоненты были бессмертны, считать их живыми не приходится. Геном человека, полный набор ДНК в каждой клетке, также не бессмертен, поскольку он постоянно меняется в процессе, называемом мейотической рекомбинацией, который сопровождает половое воспроизведение, когда одна цепочка — генов — заменяется другой (обсуждение этой проблемы также содержится в главе 2). Тут мы меняем уровень, переходя к генам, репродуктивно активным цепочкам ДНК. Ген потенциально бессмертен — пока не подвергнется мутации — так как он переносится от генома к геному, от мыши к мыши, оставаясь практически неповрежденным.[3] Является ли тогда именно он единицей отбора? В своей книге Адаптация и естественный отбор (1966) Джордж Уильямс утверждал, что ген следует рассматривать как некоторую порцию хромосомного материала, потенциально сохраняющуюся в поколениях, число которых достаточно для того, чтобы считать ее единицей естественного отбора. Оксфордский зоолог Ричард Докинз (р. 1941) в своей заслуженно знаменитой книге Эгоистичный ген (1976) безжалостно развил эту идею и проследил, как посредством эгоистического действия ген разворачивается в биосистему и обеспечивает свое собственное выживание. Я упоминал в Прологе, что наука, как правило, углубляет свои прозрения и умножает свои успехи, переходя на более высокие уровни абстракции. Эта тенденция заметна и в биологии. Естественный отбор играет роль естественной кучи компоста для произрастания абстракций, а идентификация гена как единицы отбора явилась главным шагом в данном направлении. Докинз ищет поэтому естественный отбор, протекающий на самом низком уровне, в генах, и считает организм лишь временно используемым сосудом, который безжалостно эгоистичный (в техническом смысле, я подчеркиваю) ген арендует, чтобы обеспечить свое собственное распространение. Бессознательный ген формирует свой сосуд, его фенотип, так, чтобы наилучшим образом приспособить его к окружающей среде, ибо именно наиболее приспособленный сосуд будет гарантировать распространение гена. Но существует более низкий уровень отбора, еще более абстрактный, чем ген, потенциально даже более бессмертный. Ген кодирует фенотипическую информацию, такую как информация о строении тела, его окраске или физиологических модификациях, необходимых для усиления громкости рыка. Ген представляет собой физическую целостность, которая вынуждена обновляться, когда метаболические процессы копируют нити ДНК и обеспечивают поступление копий в каждую клетку и к следующему поколению. Сам по себе, как физическая целостность, даже ген не бессмертен, ибо физический ген должен непрерывно перестраиваться. Тот факт, что в ДНК закодирована информация, только деталь, ведь функция не есть основание. Но когда мы рассматриваем ген как единицу отбора, мы в действительности фокусируемся на информации, которую он передает, и точно так же как тело организма пригодно для использования в качестве сосуда для генов, так и последовательность элементов ДНК есть подходящая физическая реализация для информации, содержащейся в гене. Подлинно бессмертной компонентой жизни является не физический ген, ею является абстрактная информация, которую он содержит.[4] Информация бессмертна, и информация безжалостно эгоистична. Генетическая информация, вероятно, является предельной единицей отбора, с ДНК в качестве своей реализации и с телом в качестве сбрасываемого служебного сосуда. Живой мир возникает, когда неорганическое вещество спотыкается на пути, проходящем среди сложной, непредсказуемой информации, и обнаруживает, что может обеспечить бессмертие для этой информации, непрерывно воспроизводя ее. И здесь зарыта еще одна неистово бегущая Красная Королева, ибо постоянство достигается только вечным воспроизведением. Подобным же образом наш собственный номинально цивилизованный, культурный, разумный и рефлексирующий уровень жизни возник, когда организмы на пути, проходящем среди сложной, непредсказуемой информации, споткнулись о другие организмы, окружающие их и следующие за ними. Когда это случилось, был изобретен язык и все человеческие организмы, прошлые, настоящие и будущие, эффективно спаялись вместе в единый мегаорганизм с потенциально неограниченными возможностями. После этой риторической, но прочувствованной пышности слога самое время спуститься к проблемам пола. Одним из наиболее загадочных проявлений естественного отбора является эволюция полового воспроизводства. На первый взгляд пол выглядит хорошей идеей, в том смысле, что он наделяет виды генетической гибкостью и быстрой реакцией на меняющиеся условия. Однако здесь имеются проблемы. Во-первых, пол не является необходимым. Довольно многие виды превосходно обходятся без него. Партеногенез (непорочное зачатие) обычное дело у растений, где его более уместно называть партенокарпией. Мы уже упоминали партенокарпические одуванчики, но могли бы добавить и много других распространенных растений, таких как черная смородина (Rubus) и манжетка обыкновенная (Alchemilla). Неполовым путем размножаются и некоторые рептилии, наиболее заметны ящерицы Нового Света рода Cnemidophorus (семейство Teiidae), ящерицы Старого Света рода Lacerta (семейство Lacertidae) и слепозмейка (Raphotyphlops braminus; семейство Typhlopidae). Никакие млекопитающие неполовым путем не размножаются, если не считать противоположных утверждений из Библии. Во-вторых, пол неустойчив. Предположим, что некоторый вид размножается половым путем, и каждая пара производит многочисленное потомство, в котором половина мужских особей и половина женских. Чтобы численность популяции оставалась приблизительно постоянной, все потомство, за исключением примерно двух особей, должно погибнуть, чтобы остались в среднем одна мужская особь и одна женская. Предположим теперь, что с одной из женских особей случилась мутация, и она приобретает способность размножаться неполовым путем. Она снова произведет многочисленное потомство, из которого выживут примерно двое; однако они, являясь клонами матери, будут обе женского пола. Обе могут размножаться с помощью партеногенеза, производя еще больше самок. При условии, что одна асексуальная самка производит столько же потомков, сколько пара сексуальных партнеров (спорное предположение, разумеется, поскольку отцы часто выполняют определенные функции и после копуляции), через несколько поколений партеногенетическая женская популяция затопит исходную. Чтобы обеспечить устойчивость, у пола должно быть некое уравновешивающее преимущество. В-третьих, пол в высшей степени сложен. Половое воспроизведение зависит от загадочного механизма мейоза, в котором, как мы увидим в главе 2, число хромосом в зародышевых клетках (гаметах, сперме и яйцеклетке) делится пополам, но восстанавливается снова в соматических (типичных для тела) клетках после оплодотворения. Насколько же невероятно мощным должно быть селективное давление, чтобы привести к развитию столь «умного» механизма? Нет ничего необычного в развитии сложных механизмов путем сталкивания вместе и видоизменения уже существующих анатомических и биохимических свойств — примером являются бесчисленные независимые случаи эволюции глаза — но, как и само обладание глазом, пол был подарком, захватывающим дух, предложением, от которого организм не мог отказаться. Оксфордский биолог Уильям Гамильтон (1936-2000), которого Ричард Докинз считал кандидатом на титул самого выдающегося дарвиниста после Дарвина, полагал, что он идентифицировал этот подарок. Гамильтон глубоко интересовался паразитами, и незадолго до того, как он сам был иронически и трагически сражен одним из них, заболев малярией, предположил, что пол дает возможность организму оставаться на шаг впереди охотящихся на него паразитов. Совместная эволюция паразита и хозяина, в которой каждый из участников создает быстрое изменение среды для эволюции другого, нуждается в быстром и специфическом типе реакции, которую может обеспечить пол. Тщательный анализ динамики сосуществования, скорее похожего на маневрирование наций в период «холодной войны», показывает, что пол создает преимущество, обеспечивая механизм сохранения генетической информации, которая становится избыточной, но может быть востребована снова, когда генотип паразита возвращается к предшествующей инкарнации. Иными словами, пол создает хранилище для мечей в эпоху мушкетов, но мушкеты могут быть сняты с вооружения. Припрятанные мечи, однако, бесполезны, если мушкеты уступают место ядерному оружию; то есть пол бесполезен, если паразит скорее вырабатывает новую стратегию, чем возвращается к старой. Эта теория остается умозрительной, так как зависит от наличия специфической эволюционной связи между паразитом и хозяином, и ее трудно подтвердить экспериментально. Легче идентифицировать механизмы, которые поддерживают пол, чем механизмы, давшие начало этому сложному устройству. Во-первых, популяции с половым размножением более гибко приспосабливаются к окружающей среде, чем партеногенетические популяции. Так, благоприятные мутации могут произойти в обоих родителях по отдельности и даровать преимущество их потомкам, в партеногенезе одна мутация следует за другой. То есть в половых популяциях мутации могут происходить параллельно, а в неполовых только последовательно. Во-вторых, в половой популяции менее вероятно распространение вредных мутаций, потому что дефектные родители еще могут произвести нормального ребенка (это становится очевидным с точки зрения менделевской наследственности, глава 2), в то время как организмы, размножающиеся неполовым путем, могут избежать скверной мутации, только если тот же самый ген мутирует обратно, что невероятно. Половой диморфизм (различия во внешности самцов и самок одного и того же вида), в частности, экстравагантную внешность, которой часто отмечены самцы, тоже относительно легко объяснить или, по крайней мере, состряпать ему правдоподобное объяснение. Например, в королларии к своей теории эволюции пола Гамильтон считает, что чрезмерно пышный внешний вид самца является знаком того, что он здоров и свободен от паразитов. Исследование самца, приводимое самкой — то, что мы, люди, можем назвать «влюбленностью», — становится тогда похожим на медицинскую проверку. Пол, по-видимому, дарует преимущества на многих уровнях популяциям, индивидам и генам. Большая часть эволюционных сдвигов создает лишь малые преимущества: чтобы расплатиться за пол, преимущество должно быть огромным. И почему вообще должно возникнуть какое-то преимущество от смешивания генов кого-то вам незнакомого с вашими? Камнем преткновения здесь является то обстоятельство, что происхождение пола, например, продолжительность пути, который организмы должны пройти, чтобы до него добраться, остается тайной до сих пор. Давайте перейдем от ощущения, что Земля движется, к реально движущейся Земле. Никогда тектонические процессы не оказывали большего воздействия на наше собственное существование, чем при едва различимых переменах, которые произошли, когда африканская кора покрылась рябью, реагируя на давления, действовавшие на нее во время путешествия по южному полушарию. Около 20 миллионов лет назад поверхность Африки была по большей части плоской и покрытой на всем своем простирании тропическим лесом. Вы могли бы начать различать неоднородности лишь с момента 15 миллионов лет назад, когда локальное поднятие образовало плоскогорья из лавы, расположенные в областях, которые мы теперь называем Кенией и Эфиопией. Эти плоскогорья сформировались в чувствительной зоне, поскольку земля под ними начала разъезжаться. Когда щель между ними расширилась, плоскогорья обрушились, образовав глубокий и длинный разлом, Великую рифтовую долину, которая теперь простирается от современного Мозамбика через Эфиопию к Красному морю и далее, до самой Сирии. Вновь выросшие в результате этого возвышенности стали преградой для дождей в восточной части континента, и тропические леса постепенно выродились в открытую саванну. Теперь ландшафт создавал богатое разнообразие потенциальных сред обитания — несколько биомов — с влажными, жаркими, богатыми растительностью зонами в одних областях и сухими травянистыми пространствами в других. Для исследования оказались открытыми не только новые ниши, для исследования и освоения открылась и репродуктивная изоляция, ибо сообщества организмов были лишены возможности мигрировать через выросшие естественные барьеры. Организмы оказались в ловушке. Организмы оказались в ловушке физического, но не эволюционного пространства. Одним из важнейших следствий естественного отбора является существование Homo sapiens, вида, который сам H. sapiens считает апофеозом эволюции. Дарвин весьма чувствительно относился к выводам из своей теории, согласно которым человек скорее был потомком обезьян, чем отдельным творением. В стане христиан также были обеспокоены мыслью, что появление человека без участия грехопадения подрывает основание христианской церкви, центральной догмой которого является идея искупления первородного греха. Как бы то ни было, существуют недвусмысленные свидетельства того, что вы и я произошли от обезьяноподобных предков. Это происхождение столь важно для того, чтобы начать понимать самих себя и свое место в биосфере, что кажется уместным потратить некоторое время на его рассмотрение. В естественном отборе необходимо учитывать расселение (миграцию жизни на суше), двуногость (хождение вертикально на двух ногах, дающее рукам свободу манипуляций), энцефализацию (разрастание мозга относительно размеров тела) и возникновение культуры. И сегодня спорят, предшествовала ли двуногость расселению или следовала за ним. Одно из преимуществ двуногости в том, что она дает большую выносливость для преследования стад, и прямоходящее животное может дальше видеть через саванну и замечать хищников. Тем, кто думает, что принятие культуры — включая употребление орудий — послужило трамплином для нашего наступления, оппоненты доказывают, что первым шагом была энцефализация. Приматы обычно обитают на деревьях в тропических и субтропических лесных экосистемах. Мы распознаем их, отмечая характерные черты анатомии их рук и ног, способы их передвижения, возможности зрения, архитектуру их зубов и их интеллект. Эта последняя черта является центральной, поскольку первичной характеристикой приматов является эволюция интеллекта как образ жизни. Характеристики зубов важны, поскольку они позволяют нам установить тип диеты и, в частности, жило ли животное на деревьях, питаясь мягкими фруктами, или на земле, поедая более твердые семена и зерна. Приматы разбиты на две основные группы — полуобезьяны и антропоиды. Полуобезьяны включают лори и буш-беби; антропоиды — мелких обезьян, крупных обезьян и людей.
Рисунок 1.10 показывает генеалогическое дерево человека, и на нем стоит задержать взгляд не в последнюю очередь потому, что отдаленная человеческая история есть нечто, приводящее в замешательство и все еще требующее, чтобы его кусочки собрали вместе. Давным-давно, в эпоху Палеоцена («старую современную») Третичного периода (первоначально «третьей» эры) Кайнозойской («современной животной») эры ранние полуобезьяны разделились на современных полуобезьян, которые далее нас не касаются, и эволюционную ветвь, в будущем ведущую к нам. Во времена раннего Олигоцена (неподражаемого «мало современного») мелкие обезьяны Нового Света (платиррхины, что означает просто «с плоским носом», включающие в себя мартышек, ревунов и капуцинов) разбили лагерь в Южной Америке. Наша ветвь дерева отделилась позже, когда мелкие обезьяны Старого Света (катаррхины, что значит «с вдавленным носом», включающие живущих на деревьях толстотелов и живущих на земле макак и резусов) появились в Африке. Гоминиды впервые появились около 30 миллионов лет назад, в Миоцене («среднем современном») в форме дриопитеков (или «жителей дубов», что отражает их предполагаемое тропическое и субтропическое место обитания). Крупные обезьяны раннего Миоцена, такие как проконсул, по осанке были очень похожи на мелких обезьян: сам проконсул не имел хвоста (как крупные обезьяны), но держал тело параллельно земле (как мелкие обезьяны). Проконсул также имел противопоставленный большой палец, что является легким намеком на возможность использования им орудий. Ископаемых гоминидов находят почти во всех отложениях Миоцена в Африке и Евразии, и самыми ранними образцами является вид проконсул с возрастом в 22 миллиона лет. Примерно на половине пути через Миоцен, между 10 и 15 миллионами лет назад, дриопитеки, которых находят даже на севере вплоть до Испании и Венгрии, разветвились на несколько родов, которые включали Сивапитеков в Индии, Пакистане, а возможно, даже в Турции, и Рамапитеков в Африке. Сивапитеки, вероятно, предки орангутангов; Рамапитеков долго считали предками человека, но теперь думают, что они были просто одним из многих обезьяноподобных видов того времени. Гоминиды, жившие около 3 миллионов лет назад, разбились на две группы, одна с большим мозгом и маленькими коренными зубами, другая с маленьким мозгом и большими коренными зубами. Первая образовала род Homo; последняя — австралопитеков рода Austropithecus (южных обезьян). Первый образец последней группы, A. africanus, был обнаружен в 1924 г. при разработках карьера в Тунге, близ Йоханнесбурга, и казался предком Homo. Первоначально об этом была опубликована лишь небольшая заметка, главным образом потому, что свежие и болезненные воспоминания о пилтдаунской подделке породили скептицизм, и существовала точка зрения, что Африка неподобающее место для колыбели человечества; Англия гораздо предпочтительнее, лучше графства, прилегающие к Лондону, но юго-западные тоже сойдут. До сих пор не определено, в какое место филогенетического дерева следует поместить A. africanus. В 1962 г. Луис Лики, старейший из охотников за ископаемыми гоминидами, проводя раскопки в ущелье Олдуваи на равнине Серенгети в Танзании, наткнулся на останки гоминида, пользовавшегося орудиями, которого он определил как новый вид Homo habilis («человек умелый»), существовавший примерно 1,8 миллиона лет назад. Изготовление орудий из камня возникло, по-видимому, приблизительно в то же время, что и значительное развитие мозга, около 2,5 миллионов лет назад, и можно лишь предполагать, способствовало ли изготовление орудий росту мозга, или наоборот. Тогда H. habilis вызвал серьезные споры, и на том основании, что внутри вида бывает много физических вариаций, некоторые думали, что это большой A. africanus, в то время как другие полагали, что это маленький Homo erectus (с которым мы сейчас встретимся). Современная точка зрения, видимо, состоит в том, что H. habilis является самостоятельным видом или, по крайней мере промежуточным видом на пути к H. erectus. Современная точка зрения, кажется, состоит также в том, что несколько скелетов, соединенных вместе в виде H. habilis, представляют на самом деле два отдельных вида, H. habilus и Homo rudolfensis, последний (названный в честь озера Рудольф, где он был найден, теперь озеро Туркана в северной Кении) имел немного более крупный мозг и более современную структуру мозга. До сих пор неизвестно, какой из этих видов в конце концов привел к H. sapiens, поскольку «очевидный» ответ, что это H. rudolfensis, приходит в противоречие с наблюдениями, согласно которым он имеет не те видимые анатомические черты, которые для этого необходимы. Теперь (около 2 миллионов лет назад) и здесь (в Африке) на сцену выступает Homo ergaster («человек работающий»), обремененный комплектом своих каменных инструментов. Он выше своих предшественников и имеет более крупный мозг. Почти полный скелет «турканского мальчика» (рис. 1.11), найденный Ричардом Лики в 1984 г. на западном берегу озера Туркана является представительным: его сохранность позволяет предположить, что он, вероятно, утонул примерно 1,6 миллиона лет назад, поскольку не был разорван на куски плотоядными животными. Homo ergaster осваивал широкие открытые пространства и умеренный сухой климат; он пользовался орудиями, которые классифицируются как Ашельские. Это название происходит от Сент-Ашель во Франции, где была найдена коллекция относительно усовершенствованных двусторонних каменных орудий, таких как каплеобразный ручной топор. Предполагают, что H. ergaster был скорее охотником, чем собирателем.
Около 1,8 миллиона лет назад H. ergaster распространился в Азию, и здесь, как и в Африке, эволюционировал в H. erectus («человек прямоходящий»), который уже полностью ходил вертикально на двух ногах и был, возможно, так же высок, как современные люди, но с более тяжелым лицом, низким лбом и надбровными дугами (рис. 1.12). Предполагается, что H. erectus следовал за стадами, которые, в свою очередь, следовали за отступлением лесов, вызванным ледниковым охлаждением планеты, и сопровождал их через тропические и субтропические зоны Саудовской Аравии до юга Центрального Китая. Отсюда он перебрался по перешейку на Яву, где в 1891 г. Эжен Дюбуа в ходе своих поисков «пропущенного звена» нашел на реке Соло части «яванского человека».
В соответствии с «африканской гипотезой» H. sapiens («человек разумный»), по-видимому, возник в Африке 150 тысяч лет назад, возможно, от H. ergaster, но, возможно, и от не столь яркого (с точки зрения скудости свидетельств о размерах его мозга) H. erectus. В свою очередь, H. sapiens пришлось конкурировать с H. neanderthalensis, образцом «пещерного человека», чьи останки нашли рабочие каменоломен в 1856 г. в Фельдерхофер Гротто над рекой Дюссель в долине Неандер в Германии (рис. 1.13).
Неандертальцы имели более крупные тела, были более мускулистыми, более коренастыми и с более бочкообразной грудной клеткой, чем мы, и хотя их мозг был в среднем крупнее, чем мозг современного человека, в их черепах, как полагают некоторые комментаторы, ощущался недостаток фронтальных долей. Они проявили способность адаптироваться к холодному климату. Позднее их образцы находили на Ближнем Востоке, в Азии и Северной Африке. Их технология обработки камня сегодня классифицируется как Мустьерская, по имени местоположения пещер в Ле Мустье во Франции. В дальнейшем, возможно, в результате контактов с H. sapiens около 40 тысяч лет назад они заимствовали идеи из Ашельской технологии и усовершенствовали ее до версии, известной как Шательперронская, по имени соответствующего района пещер. Орнаменты, сделанные из оленьих рогов и костей, позволяют сделать предположение о том, что между этими двумя видами имела место торговля. Проблеском, осветившим домашний быт неандертальцев, является находка флейты, датируемой временем от 43 до 67 тысяч лет назад, и, возможно, неандертальского происхождения, с отверстиями, соответствующими диатонической шкале из семи нот в современной человеческой музыке. Помимо музыкальных вечеров неандертальцы имели развитую социальную организацию, сотрудничали на охоте, возможно, общались с помощью языка (хотя строение их гортани указывает на то, что их язык страдал недостатком артикуляции) и хоронили своих покойников, что можно интерпретировать как чувствительность (но можно и как обрушение свода). С другой стороны, они могли быть каннибалами — что можно было бы также объяснить чувствительностью, буквальным принятием в себя тех, кого любишь, — если отбросить наше культивируемое отвращение. Если отдельный предмет искусства, одинокий отполированный и продырявленный зуб мамонтенка, является той отправной точкой, из которой могут быть сделаны надежные антропологические экстраполяции, то неандертальцы были артистичны. Артистичностью они обладали, а вот жизнестойкостью нет, ибо H. neanderthalensis исчезли около 30 тысяч лет назад, став тупиком эволюции, сухим листом на ветке дерева жизни. Вымирание неандертальцев началось около 40 тысяч лет назад с волны вторжения, которая хлынула через Европу с востока на запад и закончилась около 27 тысяч лет назад. Завоевателями были кроманьонцы (рис. 1.14), разновидность H. sapiens, почти идентичная современному человеку и названная по имени места Кро Маньон в центральной Франции (близ деревни Лес Эйзи в Дордони), где в 1868 г. были найдены первые образцы. Кроманьонец был адаптирован к теплу, и это подтверждает точку зрения, что он (вместе со своей «ею», разумеется) прибыл из Африки. Кроманьонцы сметали все перед собой, возможно благодаря своему значительному превосходству в орудиях (т.е. в оружии), известных как Ориньякские, которые включали кость и олений рог, а также изысканную глиняную посуду, или благодаря тому, что у неандертальцев не было защиты от принесенных ими болезней, или потому, что из-за недостаточно артикулированной речи неандертальцы не могли достаточно эффективно общаться, чтобы координировать свою оборону. Кроманьонцы жили в шалашах и употребляли ловушки для животных, луки и стрелы, черенки и рукоятки для ножей и битум для их крепления. Война теперь вступила на путь индустриализации, и этому суждено было стать проклятьем человеческой истории на все последующие тысячелетия.
Современные люди, однако, когда не заняты бойней, имеют теперь все необходимое, чтобы поразмышлять о своей окружающей среде, о своей физической и психологической природе, о строении вещества, которое их окружает и которое они медленно познают, чтобы подчинить своей воле. Что будет дальше, зависит от того, куда приведут эти начинания. 1. Построенный из дерева гофер, ковчег был 300 локтей в длину, 50 локтей в ширину и 30 локтей в высоту; локоть — длина руки от локтевого сгиба до кончиков пальцев, около 45 сантиметров). 2. «Бигль» имел 27 метров в длину и 7 метров в ширину в средней части судна, что примерно эквивалентно 15×4 «дарвинов». 3. Я говорю «практически неповрежденным», потому что даже если во время мейоза, происходящего в середине гена, возникнет случайная поломка ДНК, шаг рекомбинации восстановит этот ген в новом геноме. 4. Не совсем так, энтропия в конце концов уничтожает и информацию. — Прим. пер.
ТегиПохожее
|

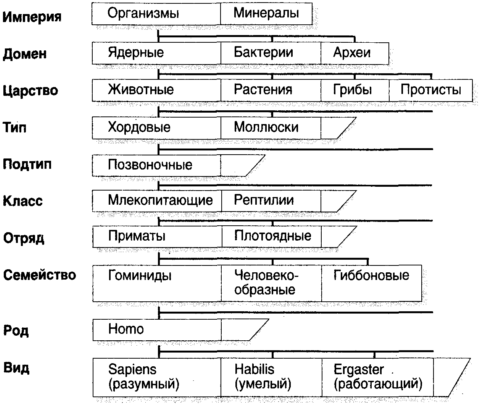
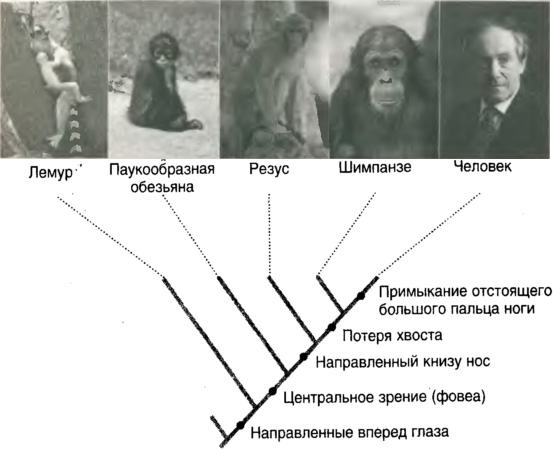
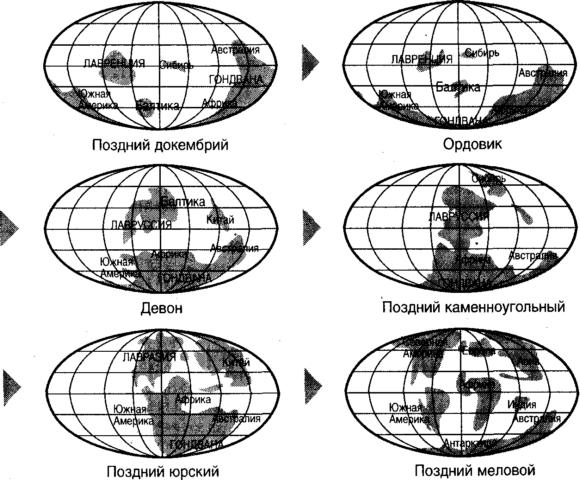

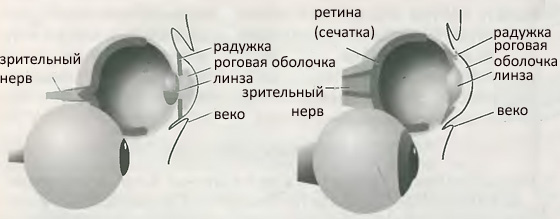
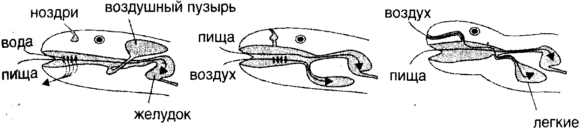
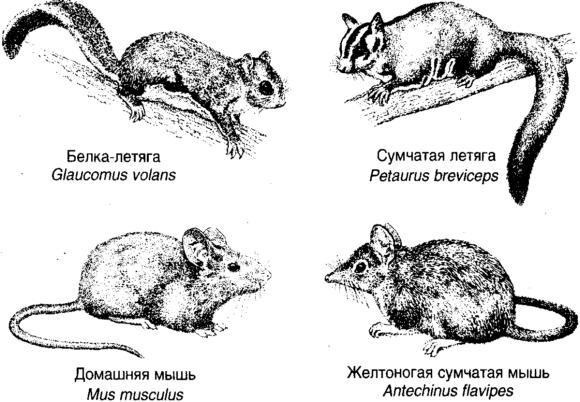

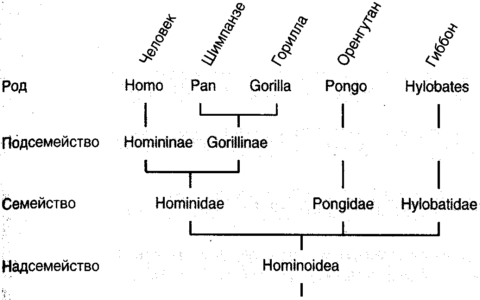
 Рис. 1.11. Несчастье индивида может быть большим подарком для науки. Это почти полный скелет Турканского мальчика (более формально WT-15000), образец H. ergaster, который без вести пропал, предположительно утонул, около 1,6 миллиона лет назад и был обнаружен в 1984 г. Важный вывод — кто-то может счесть его умозрительным — может быть сделан из деталей скелета. Относительно тяжелые кости (по сравнению с современными людьми) предполагают регулярное физическое напряжение; канал в позвоночнике, через который проходит спинной мозг, имеет меньший диаметр, чем у современных людей, что предполагает меньший поток нервных импульсов, что, в свою очередь, предполагает меньший контроль над дыханием и поэтому умозрительно отсутствие разговорной речи. Размер таза и его последствия для вынашивания плода женщинами также предполагают, что для этого вида были характерны беспомощность детей и, как ее следствие, социальная организация.
Рис. 1.11. Несчастье индивида может быть большим подарком для науки. Это почти полный скелет Турканского мальчика (более формально WT-15000), образец H. ergaster, который без вести пропал, предположительно утонул, около 1,6 миллиона лет назад и был обнаружен в 1984 г. Важный вывод — кто-то может счесть его умозрительным — может быть сделан из деталей скелета. Относительно тяжелые кости (по сравнению с современными людьми) предполагают регулярное физическое напряжение; канал в позвоночнике, через который проходит спинной мозг, имеет меньший диаметр, чем у современных людей, что предполагает меньший поток нервных импульсов, что, в свою очередь, предполагает меньший контроль над дыханием и поэтому умозрительно отсутствие разговорной речи. Размер таза и его последствия для вынашивания плода женщинами также предполагают, что для этого вида были характерны беспомощность детей и, как ее следствие, социальная организация.
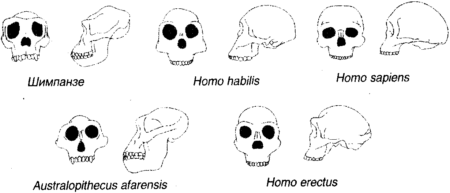

 Целью этой книги является строгое определение чисел, многочленов и алгебраических дробей и обоснование их свойств, уже известных из школы, а не ознакомление читателя с новыми свойствами. Поэтому читатель не найдет здесь новых для него фактов (за исключением, быть может, некоторых свойств, действительных и комплексных чисел), но узнает, как доказываются вещи, хорошо ему известные, начиная с «дважды два — четыре» и кончая правилами действий с многочленами и алгебраическими дробями. Зато читатель познакомится с рядом общих понятий, играющих в алгебре основную роль.
Целью этой книги является строгое определение чисел, многочленов и алгебраических дробей и обоснование их свойств, уже известных из школы, а не ознакомление читателя с новыми свойствами. Поэтому читатель не найдет здесь новых для него фактов (за исключением, быть может, некоторых свойств, действительных и комплексных чисел), но узнает, как доказываются вещи, хорошо ему известные, начиная с «дважды два — четыре» и кончая правилами действий с многочленами и алгебраическими дробями. Зато читатель познакомится с рядом общих понятий, играющих в алгебре основную роль. Знаменитая Теорема Гёделя о неполноте имеет две версии — синтаксическую (объявленную и доказанную самим Гёделем) и семантическую (чаще всего фигурирующую в популярных рассуждениях о великой Теореме). Семантическая версия утверждает, что какую бы систему формальных доказательств ни придумать, в языке найдутся истинные утверждения, не доказуемые в рамках предложенной системы. Таким образом, семантическая версия исходит из того, что некоторые выражения языка выражают осмысленные утверждения, являющиеся истинными или ложными. Синтаксическая версия не опирается на то, что какие бы то ни было выражения языка имеют какой-то смысл, она смотрит на выражения как на синтаксические конструкции, то есть как на цепочки символов, организованные по определённым правилам.
Знаменитая Теорема Гёделя о неполноте имеет две версии — синтаксическую (объявленную и доказанную самим Гёделем) и семантическую (чаще всего фигурирующую в популярных рассуждениях о великой Теореме). Семантическая версия утверждает, что какую бы систему формальных доказательств ни придумать, в языке найдутся истинные утверждения, не доказуемые в рамках предложенной системы. Таким образом, семантическая версия исходит из того, что некоторые выражения языка выражают осмысленные утверждения, являющиеся истинными или ложными. Синтаксическая версия не опирается на то, что какие бы то ни было выражения языка имеют какой-то смысл, она смотрит на выражения как на синтаксические конструкции, то есть как на цепочки символов, организованные по определённым правилам. Эрвин Рудольф Йозеф Александр Шредингер - австрийский физик-теоретик, лауреат Нобелевской премии по физике. Один из разработчиков квантовой механики и волновой теории материи. В 1945 г. Шредингер пишет книгу "Что такое жизнь с точки зрения физики?", оказавшую существенное влияние на развитие биофизики и молекулярной биологии. В этой книге внимательно рассмотрено несколько важнейших проблем. Основополагающим является вопрос: "Как могут физика и химия объяснить те явления в пространстве и времени, которые имеют место внутри живого организма?" Прочтение этой книги даст не только обширный теоретический материал, но и заставит задуматься над тем, что же в сущности есть жизнь?
Эрвин Рудольф Йозеф Александр Шредингер - австрийский физик-теоретик, лауреат Нобелевской премии по физике. Один из разработчиков квантовой механики и волновой теории материи. В 1945 г. Шредингер пишет книгу "Что такое жизнь с точки зрения физики?", оказавшую существенное влияние на развитие биофизики и молекулярной биологии. В этой книге внимательно рассмотрено несколько важнейших проблем. Основополагающим является вопрос: "Как могут физика и химия объяснить те явления в пространстве и времени, которые имеют место внутри живого организма?" Прочтение этой книги даст не только обширный теоретический материал, но и заставит задуматься над тем, что же в сущности есть жизнь? В отличие от метрической теории алгоритмов, дескриптивная теория не занимается измерением ресурсов (таких как время, объём памяти), затрачиваемых при применении алгоритма к его возможным исходным данным (в другой терминологии — к его входам). Её интересует лишь, возможен алгоритм для решения данной задачи или нет. Начальные понятия дескриптивной теории алгоритмов суть: конструктивный обьект, алгоритм, число шагов алгоритма, вычислимая функция, перечислимое множество, разрешимое множество, сводимость нумераций, главная вычислимая нумерация, вычислимая операция.
В отличие от метрической теории алгоритмов, дескриптивная теория не занимается измерением ресурсов (таких как время, объём памяти), затрачиваемых при применении алгоритма к его возможным исходным данным (в другой терминологии — к его входам). Её интересует лишь, возможен алгоритм для решения данной задачи или нет. Начальные понятия дескриптивной теории алгоритмов суть: конструктивный обьект, алгоритм, число шагов алгоритма, вычислимая функция, перечислимое множество, разрешимое множество, сводимость нумераций, главная вычислимая нумерация, вычислимая операция. Теорема Гёделя, наряду с открытием теории относительности, квантовой механики и ДНК, обычно рассматривается как крупнейшее научное достижение ХХ века. Почему? В чем ее суть? Каково ее значение? Эти вопросы в своей лекции раскрывает Алексей Брониславович Сосинский, математик, профессор Независимого московского университета, офицер Ордена академических пальм Французской Республики, лауреат премии Правительства РФ в области образования 2012 года. В частности, были даны несколько разных ее формулировок, описаны три подхода к ее доказательству (Колмогорова, Чейтина и самого Гёделя), и объяснено ее значение для математики, физики, компьютерной науки и философии.
Теорема Гёделя, наряду с открытием теории относительности, квантовой механики и ДНК, обычно рассматривается как крупнейшее научное достижение ХХ века. Почему? В чем ее суть? Каково ее значение? Эти вопросы в своей лекции раскрывает Алексей Брониславович Сосинский, математик, профессор Независимого московского университета, офицер Ордена академических пальм Французской Республики, лауреат премии Правительства РФ в области образования 2012 года. В частности, были даны несколько разных ее формулировок, описаны три подхода к ее доказательству (Колмогорова, Чейтина и самого Гёделя), и объяснено ее значение для математики, физики, компьютерной науки и философии. Всякая надежда на создание единой математической теории, амбициозного проекта, который был предложен математиком Давидом Гильбертом в 19 веке и продолжил существовать, поддерживаемый многими, в 20 столетии, рухнула. Основы математики были далеко не столь надежными, как того хотел бы Гильберт. А Гëдель своими теоремами ясно продемонстрировал, что любая система аксиом, какой бы обширной она ни была, уязвима для возникновения невосполнимых пробелов. Попытки же восполнить их созданием более полной системы породили бы только бóльшее количество утверждений без доказательств — так что и тут возникнет необходимость в усовершенствовании системы, и так далее до бесконечности. И случилось нечто странное: математики решили не обращать на это внимания. Они посчитали, что неполнота систем не имеет непосредственного влияния на их работу.
Всякая надежда на создание единой математической теории, амбициозного проекта, который был предложен математиком Давидом Гильбертом в 19 веке и продолжил существовать, поддерживаемый многими, в 20 столетии, рухнула. Основы математики были далеко не столь надежными, как того хотел бы Гильберт. А Гëдель своими теоремами ясно продемонстрировал, что любая система аксиом, какой бы обширной она ни была, уязвима для возникновения невосполнимых пробелов. Попытки же восполнить их созданием более полной системы породили бы только бóльшее количество утверждений без доказательств — так что и тут возникнет необходимость в усовершенствовании системы, и так далее до бесконечности. И случилось нечто странное: математики решили не обращать на это внимания. Они посчитали, что неполнота систем не имеет непосредственного влияния на их работу. В научных представлениях о происхождении жизни в последнее десятилетие происходит настоящая революция, и она далеко не завершена. К сожалению, эта информация доступна в основном только на английском языке. Цикл статей, предлагаемый вниманию читателей, отчасти восполнит этот пробел.
В научных представлениях о происхождении жизни в последнее десятилетие происходит настоящая революция, и она далеко не завершена. К сожалению, эта информация доступна в основном только на английском языке. Цикл статей, предлагаемый вниманию читателей, отчасти восполнит этот пробел.| Главная ≫ Инфотека ≫ Разное ≫ Книги ≫ Глава первая. Эволюция. Возникновение сложности / Десять великих идей науки. Как устроен наш мир // Питер Эткинз |
|
[time: 16 ms; queries: 7]
13 Июн 2026 12:08:04 GMT+3 |