|
||
|
|
||
| Главная ≫ Инфотека ≫ Математика ≫ Окраска глазчатых ящериц порождается клеточным автоматом // Татьяна Романовская |
Окраска глазчатых ящериц порождается клеточным автоматомТатьяна Романовская
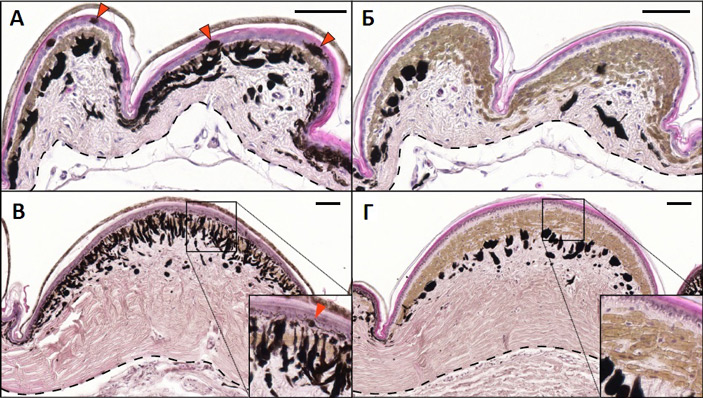
Окраска многих животных устроена причудливо и замысловато. На клеточном уровне ее возникновение описывается реакционно-диффузными моделями при помощи систем дифференциальных уравнений. В недавней работе группа ученых из Швейцарии детально изучила механизм формирования окраски глазчатых ящериц Timon lepidus. Оказалось, что это происходит по правилам, характерным для дискретного клеточного автомата, где в роли ячеек автомата выступают отдельные чешуйки кожи ящериц. Математическое моделирование позволило понять, что реакционно-диффузная система может порождать клеточный автомат благодаря особым условиям — в данном случае это подходящие размеры чешуек и толщина кожи ящериц внутри и на границе чешуек. Раскраска кожи позвоночных в макроскопическом масштабе (видимая невооруженным глазом) часто выглядит причудливо, образуя замысловатые узоры в виде полос, пятен, завитков, лабиринтов и т. д. Она может быть сформирована с самого рождения, а может меняться на протяжении жизни животного. Формирование окраски зависит от взаимодействия клеток как на ближних, так и на дальних расстояниях, возникающем при прямых контактах клеточных мембран или при помощи растворимых сигналов, которые распространяются в межклеточной среде. Рептилии (наряду с рыбами) — удобные модельные объекты для изучения механизмов формирования окраски кожи, поскольку демонстрируют широкое разнообразие рисунков и расцветок. Эти расцветки порождаются комбинацией нескольких типов клеток. Клетки-хроматофоры содержат пигменты: черно-коричневый меланин в меланофорах, желтые птеридины/каротиноиды в ксантофорах и красные птеридины/каротиноиды в эритрофорах. Также имеются клетки-иридофоры, которые содержат упорядоченные решетки нанокристаллов гуанина и формируют структурный цвет, возникающий из-за интерференции. Распределение окраски на коже зависит от того, как сочетаются хроматофоры и иридофоры в различных ее участках. Глазчатые ящерицы (Timon lepidus) заинтересовали ученых в связи с тем, что рисунок у них на спине меняется на протяжении жизни. Сразу после вылупления из яиц эти ящерицы коричневые с несколькими десятками белых пятен (рис. 1). Позднее большинство чешуек приобретает зеленый цвет, а некоторые становятся черными (в основном те, которые изначально были локализованы по периферии белых пятен). В ходе дальнейшего развития чешуйки иногда переключаются между двумя состояниями, меняя цвет с зеленого на черный или наоборот. В результате на спине ящерицы формируется рисунок в виде зеленого лабиринта на черном фоне. Особого внимания заслуживает тот факт, что в пределах одной чешуйки (на спине) у этого вида ящериц никогда не сочетаются участки разного цвета. Чешуйка может быть только полностью зеленой или полностью черной. То есть по всей площади чешуйки распределение хроматофоров абсолютно равномерно, а в соседних чешуйках либо такое же (если цвет тот же), либо отличающееся (если цвет другой). На рис. 2 показано гистологическое строение чешуек молодых и взрослых животных в поперечном разрезе.
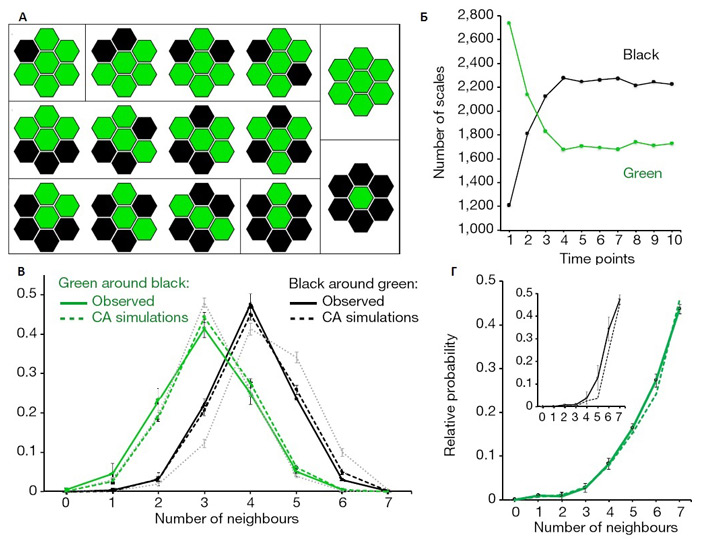
Каков же внутренний механизм создания такого необычного узора и по каким правилам происходит смена цветов чешуек? Именно этот вопрос и оказался в центре внимания исследователей, среди которых был и лауреат Филдсовской премии математик Станислав Смирнов. Авторы следили за переходом от ювенильной к взрослой окраске, чтобы выяснить механизм, определяющий дискретность окраски чешуек взрослых ящериц. Ученые фотографировали животных и обсчитывали характеристики распределения пигментов, начиная с двухнедельного возраста и до 3–4 лет с перерывами от нескольких недель до трех месяцев — всего в 10 временных точках. Учитывались тысячи чешуек на каждом животном! По мере роста ящерицы количество и взаиморасположение чешуек не изменяется, каждая чешуйка лишь увеличивается в размере. Это позволило ученым отследить «цветовую историю» каждой отдельной чешуйки. За время наблюдений (до 4 лет) сменить цвет успевали около 1500 чешуек на каждой ящерице. Для описания формируемого чешуйками рисунка авторы работы применили модель вероятностного клеточного автомата. У чешуек шестиугольная форма и к каждой примыкает шесть других чешуек, каждая из которых имеет зеленый или черный цвет. Модель предполагает, что вероятность приобретения чешуйкой того или иного цвета зависит от того, сколько других чешуек рядом с ней имеют такой же цвет. Данные, полученные в модели, очень хорошо согласовывались с тем, как менялся цвет чешуек у ящериц, что видно на рис. 3. В среднем у взрослой особи вокруг каждой зеленой чешуйки располагаются четыре черных и две зеленых чешуйки. Вероятность смены цвета чешуйки с зеленого или черного цвета на альтернативный цвет в каждой временной точке определенным образом зависит от того, сколько чешуек такого же цвета примыкало к данной чешуйке в предыдущей временной точке. Смена цвета чешуек продолжается на протяжении всей жизни животного, хотя постепенно это происходит все реже, и это снижение частоты смены цвета чешуек также согласовывалось с предсказаниями модели клеточного автомата, работающего по установленным правилам.
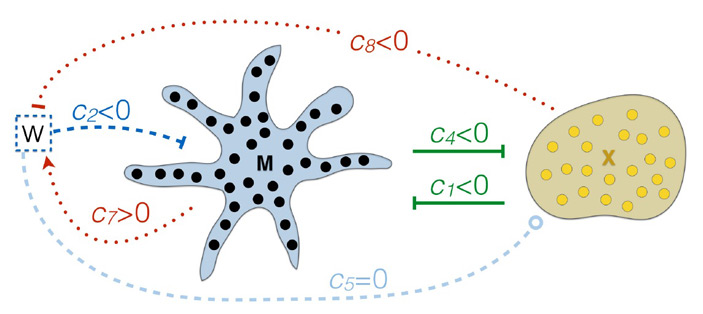
Распределение пигментных клеток в коже животных определяется взаимодействием между хроматофорами на больших и малых расстояниях. Часть этих взаимодействий активирующие, часть — ингибирующие (рис. 4). Клетки реагируют на получаемые сигналы апоптозом, миграцией или дифференцировкой, а также выделением собственных сигналов.
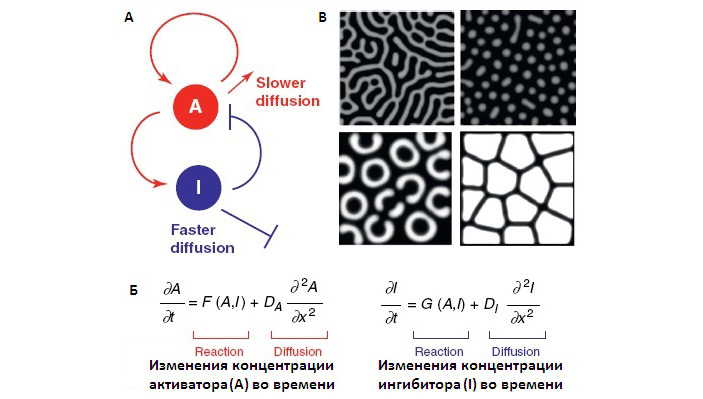
Дальнейшая работа была нацелена на то, чтобы понять, каким образом из системы этих взаимодействий рождается наблюдаемое распределение окраски на спине глазчатой ящерицы. А именно: На основе известной структуры взаимодействий между хроматофорами можно построить систему дифференциальных уравнений, описывающих поведение этой системы в рамках реакционно-диффузной модели Тьюринга (см. Reaction-diffusion system). Эта структура взаимодействий (показанная на рис. 4) и система уравнений были установлены ранее, в ходе изучения механизмов формирования полосатого рисунка на брюшке рыбок Danio rerio. Авторы использовали эти данные и уравнения для анализа окраски кожи глазчатой ящерицы. Модель Тьюринга объясняет формирование узоров изменениями локальных концентраций сигналов. Локальная концентрация сигнала описывается уравнением реакции-диффузии и зависит от соотношения скоростей появления, деградации и диффузии сигнала. Неравномерное распределение сигнала в пространстве обуславливается системой активирующих и ингибирующих взаимодействий двух (или более) продуктов реакции, которые распространяются (диффундируют) с разной скоростью. Распределение будет давать различные рисунки при различных соотношениях констант в уравнениях реакции-диффузии (как на рис. 5). Эта модель многократно доказала свою состоятельность при объяснении морфогенеза при развитии живых организмов. В роли активаторов и ингибиторов в этом случае выступают сигнальные молекулы, заякоренные в мембранах клеток (контактные взаимодействия ближнего действия) или диффундирующие молекулы-морфогены (сигналы дальнего действия).
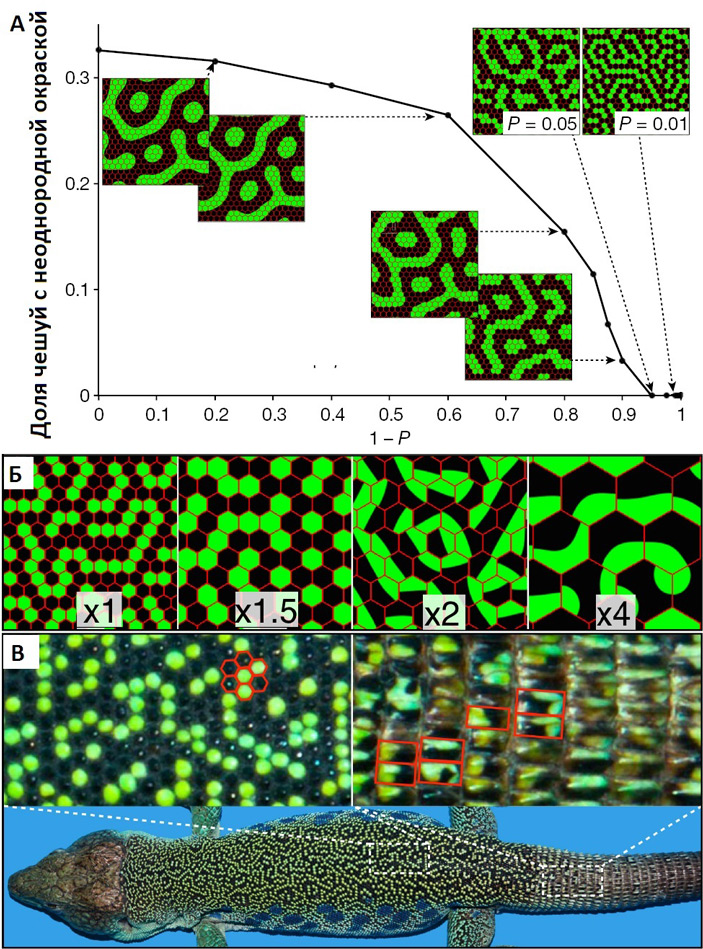
Авторы выдвинули и проверили при помощи математического моделирования предположение о том, что распределение окраски чешуек определялось геометрией тканей кожи и было следствием разницы скоростей передачи сигналов в пределах одной чешуйки и на границе между чешуйками: во-первых, из-за различий толщины кожи, во-вторых, из-за сниженного числа хроматофорных клеток на краю чешуек и в разделительной области (это хорошо видно на рис. 2). Исходя из этих двух условий, можно предположить, что в области границы будет существенно снижаться абсолютное и относительное количество сигнала, передаваемого посредством контактного взаимодействия хроматофоров. Эти факторы были учтены посредством внесения соответствующих поправок в систему уравнений реакции-диффузии. В результате при определенных значениях коэффициентов в модели возникала дискретная раскраска чешуек (рис. 6, А), причем эта дискретность нарушалась, если площадь чешуек превышала некоторое пороговое значение. Нужно заметить, что более крупные прямоугольные чешуйки на хвосте глазчатых ящериц, которые имеют значительно больший размер, чем чешуйки на коже спины, действительно утрачивают дискретную окраску, — некоторые хвостовые чешуйки окрашены частично в черный, а частично в зеленый цвет (рис. 6, Б, В).
Таким образом, в работе на новом модельном объекте (глазчатой ящерице) было установлено, что сочетание реакционно-диффузной модели Тьюринга с неоднородностью среды (геометрическими параметрами тканей) может порождать поведение системы, характерное для клеточного автомата. Можно также отметить, что само распределение чешуек в коже может быть также порождением некой системы отношений реакции-диффузии, а это значит, что клеточный автомат может быть результатом суперпозиции двух систем реакции-диффузии, участвующих в формировании тканей кожи глазчатой ящерицы. Вполне ожидаемо, что эта схема может иметь место и в других случаях формирования узоров при морфогенезе живых организмов. Источник: Liana Manukyan, Sophie A. Montandon, Anamarija Fofonjka, Stanislav Smirnov & Michel C. Milinkovitch. A living mesoscopic cellular automaton made of skin scales // Nature. 2017. V. 544. P. 173–179. DOI: 10.1038/nature22031. См. также: Татьяна Романовская
ТегиПохожее
|


 Возьмите простое решетчатое пространство. Задайте набор нехитрых правил. Запустите время. Вы получили клеточный автомат — почти что целый мир.
Возьмите простое решетчатое пространство. Задайте набор нехитрых правил. Запустите время. Вы получили клеточный автомат — почти что целый мир. Сколько нужно вопросов (с ответом “да” и “нет”), чтобы заведомо отгадать задуманное число от 1 до 1000? Можно ли обойтись меньшим числом вопросов? Если нет, то как это доказать? Сколько нужно взвешиваний на чашечных весах без гирь, чтобы наверняка выделить более лёгкую монету среди 1000 одинаковых на вид? С такого рода вопросов начинается наука о сложности алгоритмов, и очень скоро доходит до важных, но до сих пор не решённых задач.
Сколько нужно вопросов (с ответом “да” и “нет”), чтобы заведомо отгадать задуманное число от 1 до 1000? Можно ли обойтись меньшим числом вопросов? Если нет, то как это доказать? Сколько нужно взвешиваний на чашечных весах без гирь, чтобы наверняка выделить более лёгкую монету среди 1000 одинаковых на вид? С такого рода вопросов начинается наука о сложности алгоритмов, и очень скоро доходит до важных, но до сих пор не решённых задач. В научных представлениях о происхождении жизни в последнее десятилетие происходит настоящая революция, и она далеко не завершена. К сожалению, эта информация доступна в основном только на английском языке. Цикл статей, предлагаемый вниманию читателей, отчасти восполнит этот пробел.
В научных представлениях о происхождении жизни в последнее десятилетие происходит настоящая революция, и она далеко не завершена. К сожалению, эта информация доступна в основном только на английском языке. Цикл статей, предлагаемый вниманию читателей, отчасти восполнит этот пробел. Продолжительность циклов большинства цикад не случайна, а представляет собой интервалы из простых чисел (чисел, делимых без остатка только на себя — 3, 5, 7, 11, 13, 17 и т. д.), являясь наиболее действенной стратегией выживания и размножения.
Продолжительность циклов большинства цикад не случайна, а представляет собой интервалы из простых чисел (чисел, делимых без остатка только на себя — 3, 5, 7, 11, 13, 17 и т. д.), являясь наиболее действенной стратегией выживания и размножения. Закон квадрата — куба представляет собой следующий принцип: если объект пропорционально (то есть с помощью преобразования подобия) увеличивается (уменьшается) в размере, его новый объём будет пропорционален кубу масштабирующего коэффициента, а новая площадь его поверхности — пропорциональна квадрату масштабирующего коэффициент. Этот закон находит своё применение в технике и биомеханике.
Закон квадрата — куба представляет собой следующий принцип: если объект пропорционально (то есть с помощью преобразования подобия) увеличивается (уменьшается) в размере, его новый объём будет пропорционален кубу масштабирующего коэффициента, а новая площадь его поверхности — пропорциональна квадрату масштабирующего коэффициент. Этот закон находит своё применение в технике и биомеханике.| Главная ≫ Инфотека ≫ Математика ≫ Окраска глазчатых ящериц порождается клеточным автоматом // Татьяна Романовская |
|
[time: 16 ms; queries: 8]
21 Июл 2026 15:27:29 GMT+3 |